動物遺伝制御学研究室
Laboratory of Animal Genetics
Laboratory of Animal Genetics
脊椎動物の染色体を構成する反復DNA配列の構造と進化
脊椎動物の染色体には、染色体の一部を構成する部位特異的な反復配列が存在し、それらはそれぞれの系統で独自の進化を遂げてきました。これらの反復配列は一般に塩基の置換速度が速く、塩基配列や染色体上の分布に関する情報は、進化過程に生じた染色体やゲノム構造の変化を調べる上で良い指標となります。私たちは、これまでに哺乳類、鳥類、爬虫類などの多岐にわたる動物を用いて、染色体のヘテロクロマチン部位を構成する様々な高度反復配列をクローニングし、その構造や染色体上の分布、異なる種間での塩基配列の相同性などを調べ、各種動物綱における染色体構造の特性や反復配列の分子進化の過程を明らかにすることを試みています。
1)ハツカネズミの動原体反復配列
2)シリアンハムスターのヘテロクロマチンを構成する反復配列
2.鳥類の反復配列
1)鳥類に存在するマイクロ染色体特異的反復配列
2)鳥類のW染色体を構築するレトロトランスポゾン由来の反復配列
1)ハツカネズミの動原体反復配列
Mus属に属するハツカネズミがもつ動原体反復配列は、一般にコピー数の多いmajorサテライトDNAとコピー数の少ないminorサテライトDNAに大別することができ、それらはゲノムの数 % を占めています(図1)。minorサテライトDNAには動原体タンパクとの結合部位が含まれキネトコアの形成に重要な働きを持っています。一方、majorサテライトDNAは変異が大きく、野生のハツカネズミでは、染色体上の分布パターンやコピー数に大きな違いが見られます。私たちは、中国やロシアを中心にユーラシア大陸の野生ハツカネズミを対象として、Mus属の系統進化と生物地理学的な視点からその変異の分布を詳細に解析しています(図2)
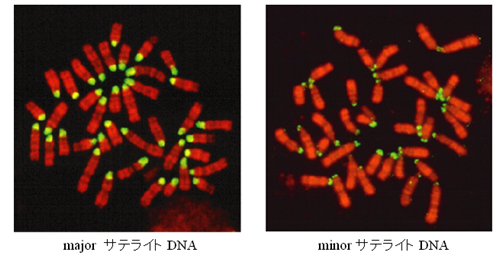
↑図1 ハツカネズミの動原体を構成する反復配列の分布パターン
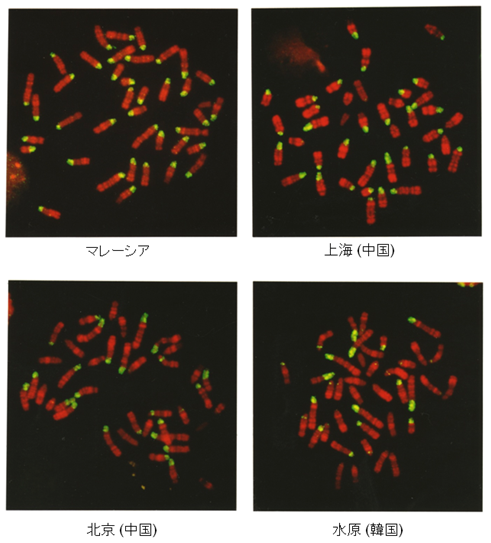
↑図2 アジアの野生ハツカネズミにおけるmajorサテライトDNAの染色体分布とコピー数の多様性
シリアンハムスターの染色体には、動原体や性染色体以外にも多くのヘテロクロマチン領域が存在します(図3)。それらは、縦列重複型のサテライトDNAや部位特異的に増幅した散在型の反復配列など多種多様な反復配列によって構成されていて、塩基の置換速度やゲノム中のコピー数も様々です。また、これらの反復配列の分子進化や染色体上の分布は、核型の進化とも密接に関係しており、脊椎動物のゲノム・染色体の進化過程とそのメカニズムを解明する上で研究モデルとなります。
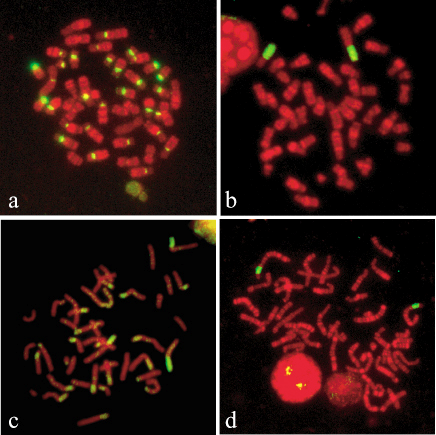
↑図3 シリアンハムスターより単離された、染色体を構成する4種の染色体部位特異的高度反復配列
(動原体領域 (a)、XY染色体 (b)、常染色体の短腕 (c)、第2染色体短腕 (d) に特異的に存在する高度反復配列のFISHパターンを示す。(c) に示した常染色体の短腕のヘテロクロマチンは、部位特異的に増幅したレトロトランスポゾン由来の反復配列によって構成されています)
1)鳥類に存在するマイクロ染色体特異的反復配列
ほとんどの鳥類の核型は、染色体サイズが大きく異なるマクロ染色体とマイクロ染色体で構成され、それらの染色体間にはサイズだけでなく、遺伝子密度、GC含量、複製時期、散在型反復配列のコピー数など、様々な構造ならびに機能的な違いが見られます。これらの結果は、鳥類の染色体にはサイズ依存的な区画化が存在することを示しています。私たちはこれまでに、数多くの鳥類からマイクロ染色体特異的な動原体反復配列をクローニングし、動原体ヘテロクロマチンの構成においてもマクロ−マイクロ染色体間で大きく異なることを明らかにしました(図4)。また、これらの反復配列は、塩基配列の置換速度が速く、種の類縁関係を示すよい指標となります。たとえば、ニワトリとニホンウズラは同じキジ科に属し41 bpを基本単位とする反復配列がマイクロ染色体の動原体部位に存在しますが、塩基配列は全く異なります(図5)。

↑図4 鳥類のマイクロ染色体の動原体ヘテロクロマチンを構築する反復配列
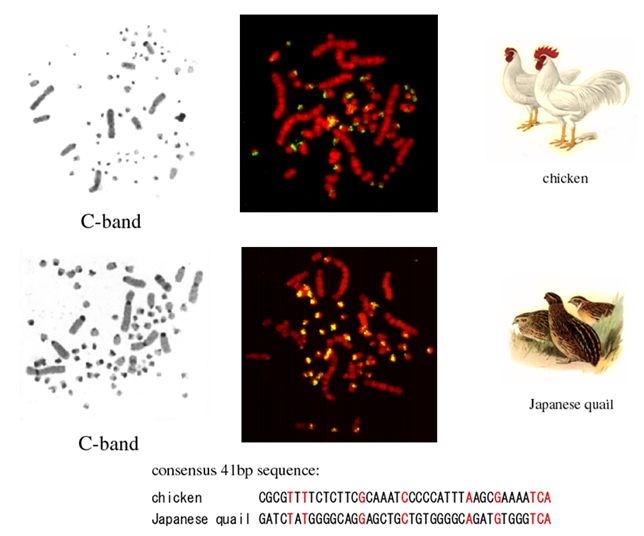
↑図5 ニワトリとニホンウズラのC染色パターンとマイクロ染色体特異的反復配列の染色体分布および塩基配列の比較
走鳥類を除くすべての鳥類では、Z-W染色体間の形態的な分化が大きく進み、W染色体の矮小化とヘテロクロマチン化がみられる。これは、W染色体に構造的な変化が生じ、Z-W染色体間で遺伝的組換えが抑制されたことによってW染色体に突然変異が蓄積し、真正染色質が失われるとともに反復配列が増幅し蓄積された結果であると考えられています。これまでに、ニワトリのW染色体のヘテロクロマチンを構成する反復配列が単離されその構造が調べられていますが、私たちは、キジ目の鳥類のW染色体でレトロトランスポゾン由来のLINE様反復配列が増幅し蓄積されていることを見出しました(図6)。この配列は、他の反復配列と異なり非常に保存性が高いことが分かっています。
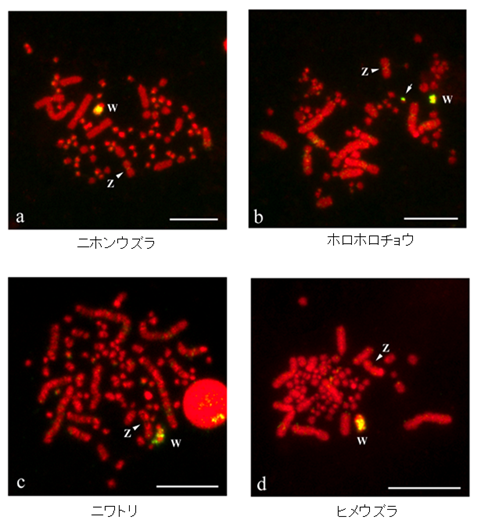
↑図6 キジ目鳥類のW染色体のヘテロクロマチンを構築するLINE (NME-Apa I 9) 様反復配列の分布パターン
同様に、爬虫類の染色体からもヘテロクロマチンを構成する反復配列が得られています。私たちは、シマヘビからヘテロクロマチンを構成する2種類のLINE配列を単離しその解析を進めています(図7)。通常ゲノム中に広く分散して存在するLINE配列が、ヘビ類では部位特異的に大量に増幅し、染色体のヘテロクロマチン領域を形成していることがわかりました。このような例は他の脊椎動物ではあまり観察されていません。

↑図7 シマヘビから得られた、染色体末端部のヘテロクロマチンを構築する2種類のLINE配列の染色体分布