動物遺伝制御学研究室
Laboratory of Animal Genetics
Laboratory of Animal Genetics
動物の種間雑種にみられる不妊や雑種退化の遺伝制御機構に関する
遺伝学的研究
遺伝学的研究
2.げっ歯類の種間雑種に見られる発育不全現象とゲノムインプリンティングによるエピジェネティックな生殖隔離機構(作成中)
.
雑種の雄性不妊現象については、Mus属のハツカネズミで最も詳しい解析がなされていますが、その遺伝的メカニズムと分子基盤については、まだ不明な点が数多く残されています。私たちは、laboratory mouse (Mus musculus domesticus)とヨーロッパ産野生ハツカネズミMus spretusの種間雑種について、細胞遺伝学的な手法を用いた詳細な解析を行いました(図1 - 3)。この種間雑種では、精子形成が精母細胞の第一減数分裂中期で停止しますが、その主たる原因が、精母細胞の減数分裂におけるX-Y染色体間の対合異常であることを明らかにしました。さらに、常染色体の対合異常も第一精母細胞の細胞死を引き起こすことを明らかにしました。現在、私たちはPhodopus属に属するドワーフハムスターの種間雑種の雄性不妊現象についても研究を進めており、F1個体における精子形成と染色体対合について詳細な解析を行った結果、様々なステージで精子形成に障害が生じることを明らかにしました(図4, 5)。さらに、種間雑種でヘテロ接合となることによって、精巣の発生が著しく抑制される雑種不妊遺伝子が存在することを見つけだし、現在、その原因遺伝子の同定を試みています(図4)。
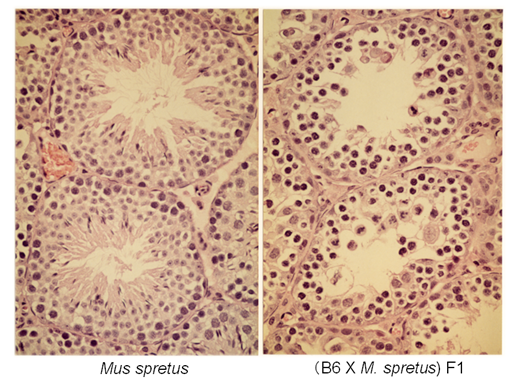
↑図1 laboratory mouse (C57BL/6系統)とMus spretus間のF1雑種の精巣組織像。精母細胞の時期で精子形成が停止していることが分かる。
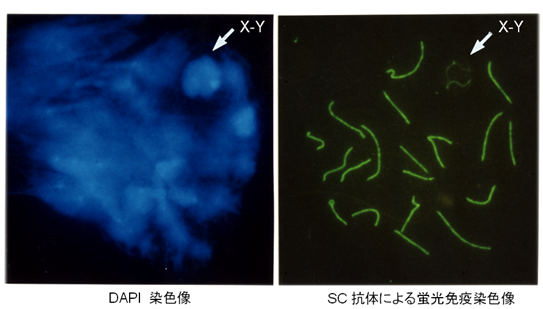
↑図2 マウスにおける第一精母細胞パキテン期の染色体像。同一細胞の核をDAPI染色とSC (Synaptonemal Complex) 抗体で蛍光免疫染色を行った。SCのコアからDNAがファイバー状に拡がっているのが分かる。XY染色体は常染色体とは異なる構造を示す。
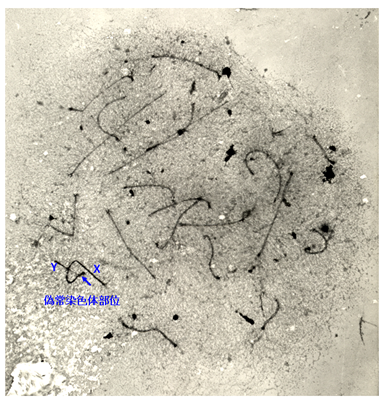
↑図3 マウス精母細胞のパキテン期における相同染色体対合の電子顕微鏡写真。銀染色によって20対の相同染色体対のシナプトネマ構造が観察できる。X-Y染色体では、常染色体とは異なり、小さな相同部(偽常染色体部位)のみで対合していることが分かる。種間雑種でX-Y染色体の対合が阻害されると、精子形成は第一減数分裂中期で停止する。
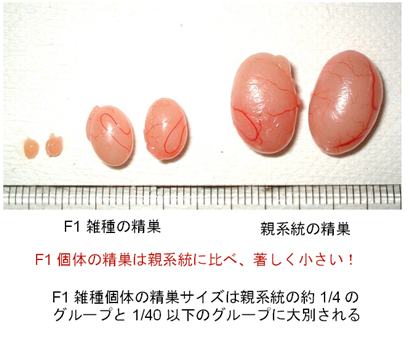
↑図4 Phodopus属ハムスターの種間雑種に観察される2つのタイプの精巣。非常に小さな精巣は、ヘテロ接合で表現型が現れる雑種不妊遺伝子に起因することが推定されている。
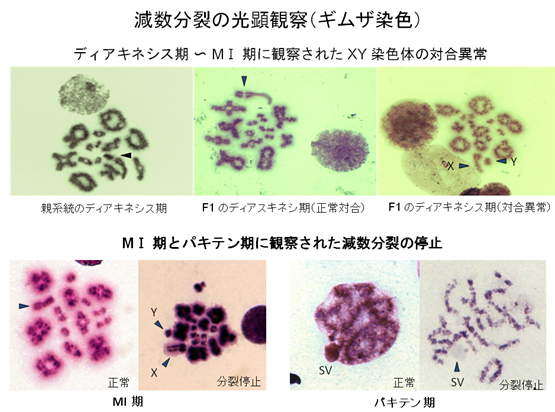
↑図5 Phodopus属ハムスターの種間雑種の第一精母細胞で観察される染色体対合のパターン。パキテン期でX-Y染色体が対合できなかった場合、第一減数分裂中期で精子形成が停止する。また、常染色体の対合異常が生じた場合は、パキテン期で減数分裂が停止する。