摦暔堚揱惂屼妛尋媶幒
Laboratory of Animal Genetics
Laboratory of Animal Genetics
愐捙摦暔偺斾妑僎僲儉妛乮Comparative Genomics乯
- 愼怓懱丒僎僲儉偺峔憿偲婡擻偐傜恑壔傪扵傞 -
- 愼怓懱丒僎僲儉偺峔憿偲婡擻偐傜恑壔傪扵傞 -
俀丏婡擻堚揱巕偺愼怓懱儅僢僺儞僌偵傛傞妀宆恑壔偺夝愅
俁丏愐捙摦暔偵傒傜傟傞僎僲儉偺儌僓僀僋峔憿偲偦偺婲尮
偝傜偵徻偟偄愢柧偼偙偪傜乮pdf僼傽僀儖乯 .
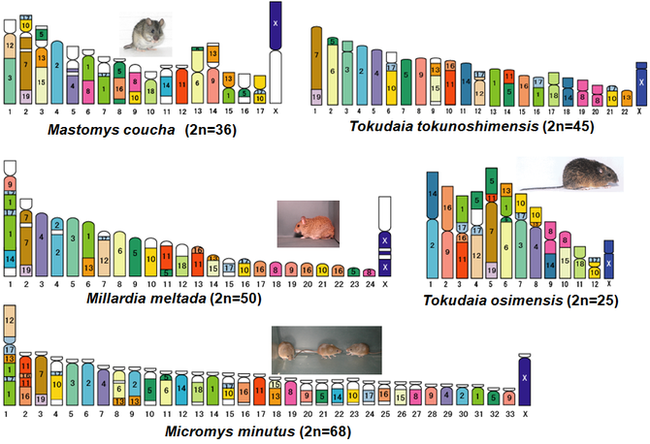
偁傞摦暔庬偐傜摼傜傟偨愼怓懱摿堎揑DNA僾儘乕僽傪寀岝昗幆偟丄暿庬偺愼怓懱偵僴僀僽儕僟僀僘偡傞偙偲偵傛偭偰丄堎側傞庬娫偱憡摨側愼怓懱椞堟傪専弌偡傞偙偲偑偱偒傑偡丅恾侾偼丄僯儚僩儕偲僉僕壢4庬偺娫偱惗偠偨峔憿曄壔傪愼怓懱儁僀儞僥傿儞僌僾儘乕僽朄偱専弌偟偨椺傪帵偟偰偄傑偡丅恾俀偼丄儅僂僗偺愼怓懱DNA傪梡偄偨愼怓懱儁僀儞僥傿儞僌偵傛偭偰嶌惉偝傟偨丄5庬偺僱僘儈垷壢摦暔偺愼怓懱抧恾偱偡丅愼怓懱忋偺僽儘僢僋偺斣崋偼丄懳墳偡傞儅僂僗偺愼怓懱斣崋傪昞偟偰偄傑偡丅偙傟傜偺寢壥傪丄宯摑庽偺忋偵僾儘僢僩偡傟偽丄恾俁偵帵偡傛偆偵丄條乆側摦暔偺恑壔夁掱偱惗偠偨愼怓懱峔憿曄壔偺僾儘僙僗傪悇掕偡傞偙偲偑偱偒傑偡丅偝傜偵丄宯摑庽傪慿偭偰偄偗偽丄婛偵愨柵偟偨慶愭偑傕偭偰偄偨妀宆傪悇掕偡傞偙偲傕壜擻偱偡丅嵟嬤偺巹偨偪偺尋媶偱丄僉僕栚偺慶愭偼丄僟僠儑僂傗僄儈儏乕側偳偺僟僠儑僂栚偺捁椶偑傕偮妀宆偲椶帡偟偨妀宆傪傕偭偰偄偨偙偲偑傢偐傝傑偟偨丅傑偨丄僱僘儈壢摦暔偺慶愭妀宆傕悇掕偡傞偙偲偑偱偒傑偟偨丅尰嵼偼丄杒奀摴戝妛棟妛尋媶堳偺惣揷愮掃巕愭惗偲偺嫟摨尋媶偱丄偝傜偵懡偔偺捁椶偵偮偄偰夝愅傪峴偄丄捁椶偵偍偗傞妀宆恑壔夁掱偺慡杄傪柧傜偐偵偡傞偙偲傪帋傒偰偄傑偡丅
仾恾侾丂僯儚僩儕愼怓懱 DNA 傪梡偄偨僉僕壢捁椶偺愼怓懱儁僀儞僥傿儞僌
仾恾俀丂僱僘儈垷壢 俆庬偵偍偗傞儅僂僗偲偺愼怓懱憡摨椞堟偺斾妑愼怓懱抧恾
仾恾俁丂儈僩僐儞僪儕傾 Cytb 堚揱巕偺墫婎攝楍偐傜嶌惉偝傟偨僉僕栚侾俁庬偺暘巕宯摑庽偲斾妑愼怓懱儁僀儞僥傿儞僌偺寢壥偐傜悇掕偝傟偨丄僉僕栚偺妀宆恑壔偺夁掱偲僉僕栚偺嫟捠慶愭偺妀宆
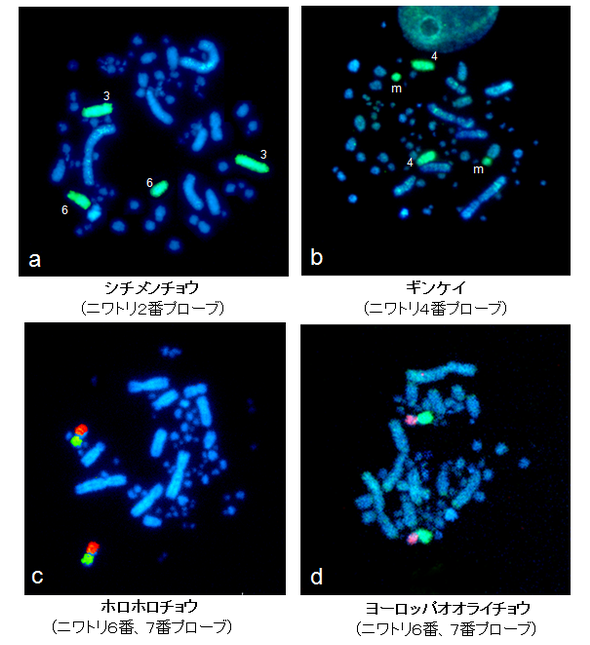
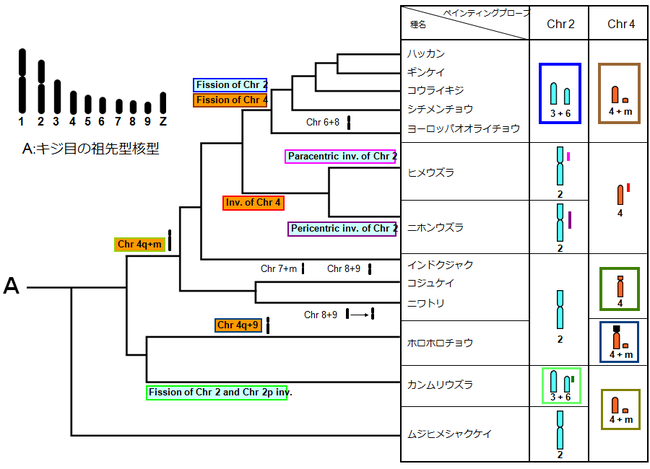
捁椶傗唳拵椶偺懡偔偼丄愼怓懱僒僀僘偑戝偒偔堎側傞儅僋儘愼怓懱偲儅僀僋儘愼怓懱偱峔惉偝傟傞摿堎揑側妀宆傪傕偭偰偄傑偡乮恾係乯丅捁椶偲唳拵椶偑栺2壄擭埲忋慜偵嫟捠慶愭偐傜暘婒偟偨偙偲傪峫偊傞偲丆捁椶偲唳拵椶娫偵傒傜傟傞妀宆偺椶帡惈偼丄愐捙摦暔偺妀宆恑壔偲惈愼怓懱偺恑壔偺夁掱傪峫嶡偡傞忋偱旕忢偵嫽枴偁傞栤戣傪採婲偟傑偡丏偟偐偟丄偙偺妀宆偺椶帡惈偑堚揱巕偺楢嵔孮偺憡摨惈傪帵偡傕偺偐丄偁傞偄偼廂澥偵傛傞嬼慠偺帠徾側偺偐偵偮偄偰偼挿偄娫晄柧偱偟偨丏捁椶偲唳拵椶偺愼怓懱偺憡摨惈傪挷傋傞応崌丄椉幰偼暘椶孮偑峧偺儗儀儖偱堎側傝丄愼怓懱傪峔惉偡傞DNA攝楍傕戝偒偔堎側傞偨傔丄僯儚僩儕偺儁僀儞僥傿儞僌僾儘乕僽傪唳拵椶偺愼怓懱夝愅偵梡偄傞偙偲偼偱偒傑偣傫丅偟偐偟丄愐捙摦暔偑嫟捠偵傕偭偰偄傞婡擻堚揱巕傪愼怓懱忋偵儅僢僺儞僌偡傟偽乮恾俆乯丄歁擕椶亅捁椶亅唳拵椶娫偱捈愙偵愼怓懱偺憡摨惈傪斾妑偡傞偙偲偑壜擻偱偡丅
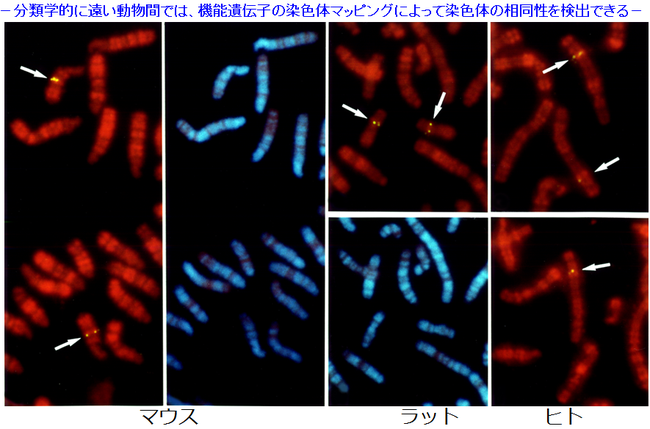
巹偨偪偼丄嫗搒戝妛棟妛尋媶壢偺垻宍惔榓攷巑偲偺嫟摨尋媶偱丄僗僢億儞偐傜摼傜傟偨戝検偺堚揱巕DNA僋儘乕儞傪僗僢億儞愼怓懱忋偵儅僢僺儞僌偟丄悽奅弶偺捁椶?唳拵椶娫偺斾妑愼怓懱抧恾偺嶌惉偵惉岟偟傑偟偨丅偦偺寢壥丄1-6斣傑偱偺儅僋儘愼怓懱偱偼丄僯儚僩儕乕僇儊娫偱1懳1偺愼怓懱偺懳墳偑傒傜傟丄僯儚僩儕偲僇儊偼栺2壄3愮擭慜偵暘婒偟偨偵傕偐偐傢傜偢丄椉幰偺愼怓懱峔憿偼傎偲傫偳曄壔偟偰偄側偄偙偲偑柧傜偐偲側傝傑偟偨丅偦偺堦椺偲偟偰丄恾俇偵僯儚僩儕偲僗僢億儞偺1斣愼怓懱偺斾妑抧恾傪帵偟傑偟偨丅偙偺恾偺恀拞偵偼僸僩愼怓懱偲偺憡摨椞堟傪昞偟偨僯儚僩儕偺堚揱楢嵔抧恾傪帵偟偰偁傝傑偡丅摨條偵僔儅僿價偵偍偄偰傕斾妑愼怓懱抧恾傪嶌惢偟丄僯儚僩儕偺愼怓懱抧恾偲斾妑偟偨偲偙傠丄僔儅僿價偱偼僯儚僩儕偲偺娫偵僗僢億儞偺傛偆側侾懳侾偺愼怓懱偺懳墳偼尒傜傟偢丄僯儚僩儕偲偺愼怓懱憡摨惈偼僇儊傛傝掅偄偙偲偑敾柧偟傑偟偨乮恾俈乯丅偙傟傜偺寢壥偼丄僇儊偺曽偑僿價傛傝傕傛傝捁椶偵嬤墢偱偁傞偲偄偆暘巕宯摑妛揑側椶墢娭學傪巟帩偟偰偄傑偡丅巹偨偪偼丄偝傜偵懡偔偺唳拵椶摦暔偵偮偄偰愼怓懱抧恾傪嶌惉偟丄捁椶偲唳拵椶偺娫偱丄偳偺傛偆側儊僇僯僘儉偱丄偦偟偰偳偺傛偆側夁掱傪宱偰愼怓懱恑壔偑惗偠偰偒偨偐傪柧傜偐偵偟偨偄偲峫偊偰偄傑偡丅傑偨丄尰懚偡傞唳拵椶偺愼怓懱偺斾妑偐傜丄婛偵愨柵偟偨嫲棾傗拞惗戙偺捁椶偺愼怓懱峔憿傪悇掕偡傞偲偄偆戝偒側柌傪幚尰偟偨偄偲巚偭偰偄傑偡丅

仾恾係丂梤枌椶偺宯摑娭學偲妀宆
仾恾俆丂僸僩丄儅僂僗丄儔僢僩偵偍偗傞 CNR8 堚揱巕偺斾妑愼怓懱儅僢僺儞僌
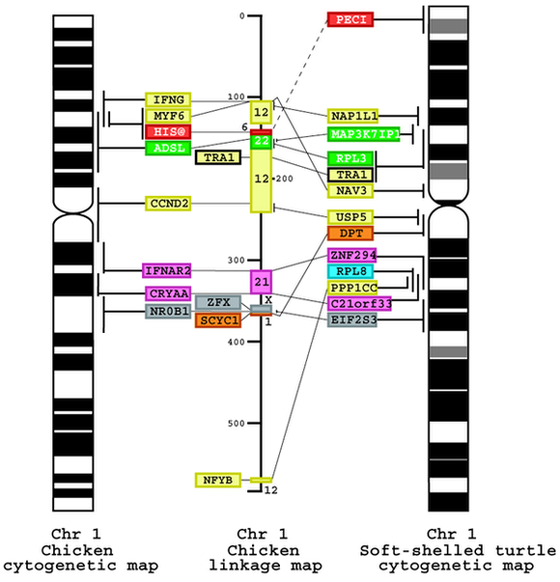
仾恾俇丂Comparative cytogenetic map of chicken and turtle chromosome 1.
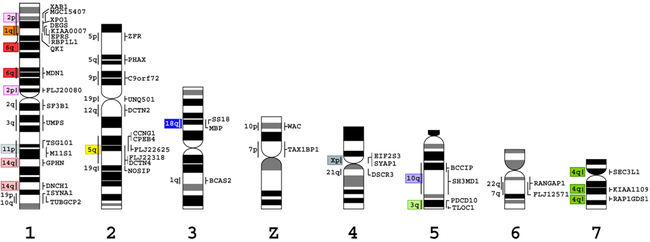
仾恾俈丂僔儅僿價偺婡擻堚揱巕愼怓懱抧恾乮僔儅僿價偺儅僋儘愼怓懱偵儅僢僾偝傟偨堚揱巕偺僯儚僩儕愼怓懱忋偺埵抲傪丄僔儅僿價愼怓懱偺嵍懁偵帵偟偰偁傝傑偡丅乯
愐捙摦暔偺僎僲儉拞偱偼ATGC偺係暥帤偑慡偔儔儞僟儉偵暲傫偱偄傞偺偐丠 捁椶偲唳拵椶偑傕偮儅僀僋儘愼怓懱偼偳偺傛偆側峔憿傪偟偰偄傞偺偐丠 偑傛偔敾偭偰偄傑偣傫偱偟偨丅偲偙傠偑嵟嬤偺尋媶偱丄歁擕椶偲捁椶偺僎僲儉偵偼丄AT-rich乮AT偑朙晉乯側椞堟偲GC?rich乮GC偑朙晉乯側椞堟偑300 kb埲忋偺僒僀僘偱儌僓僀僋忬偵懚嵼偡傞偙偲丄偦偟偰丄僯儚僩儕偱偼儅僀僋儘愼怓懱偼GC娷検偲堚揱巕枾搙偑崅偄偙偲偑柧傜偐偵側偭偰偒傑偟偨乮恾俉乯丅偟偐偟丄偙偺僎僲儉偺儌僓僀僋峔憿偑丄曄壏摦暔乮嫑椶丄椉惗椶丄唳拵椶乯偲峆壏摦暔乮捁椶丄歁擕椶乯偺暘婒揰偱惗偠偨偲偄偆愢偲丄梤枌傪傕偨側偄摦暔乮嫑椶丄椉惗椶乯偐傜梤枌傪傕偮摦暔乮唳拵椶丄捁椶丄歁擕椶乯偵恑壔偟偨帪偵惗偠偨偲偄偆愢偑偁傝傑偟偨丅偙偺撲傪夝柧偡傞尞偼丄唳拵椶偑埇偭偰偄傑偡丅偦偙偱巹偨偪偼丄棟壔妛尋媶強偺岺妝庽梞攷巑偲偺嫟摨尋媶偱丄僗僢億儞偺儅僋儘愼怓懱偲儅僀僋儘愼怓懱偵懚嵼偡傞堚揱巕娫偱GC偺娷検傪斾妑偟唳拵椶僎僲儉峔憿偺摿挜傪挷傋傑偟偨丏偦偺寢壥丄僯儚僩儕偲僗僢億儞偱偼丄儅僀僋儘愼怓懱偵懚嵼偡傞堚揱巕孮偼丄儅僋儘愼怓懱偺堚揱巕孮偵斾傋偰崅偄GC娷検傪傕偪丄愼怓懱偺僒僀僘偵埶懚偟偨GC偺嬫夋壔丄偮傑傝儅僀僋儘愼怓懱偵GC偑桳堄偵拁愊偝傟偰偄傞偙偲傪尒偮偗傑偟偨丅偙偺寢壥偼丄梤枌椶偺慶愭偱偼婛偵GC娷検偑嬫夋壔偝傟偨僎僲儉峔憿偑妉摼偝傟偰偄偨偙偲傪帵偟偰偄傑偡丏偦偟偰丄梤枌椶偵偍偗傞僎僲儉恑壔傪峫偊傞応崌丄歁擕椶偱偼屄乆偺愼怓懱忋偱GC-rich側椞堟偲GC-poor側椞堟偑摨堦愼怓懱忋偱儌僓僀僋忬偵懚嵼偡傞偺偵懳偟丄捁椶偲唳拵椶偱偼丄愼怓懱偺僒僀僘埶懚揑側GC娷検偺嬫夋壔偑惗偠偰偄傞丄偮傑傝GC-rich側晹暘偼儅僀僋儘愼怓懱偵廤栺偝傟偰偄傞偲偄偆擇偮偺儌乕僪偑懚嵼偡傞偙偲傪柧傜偐偵偟傑偟偨乮恾俋乯丅偦偟偰丄儅僂僗側偳偺陽帟椶偺宯摑偵偍偄偰偼丄僸僩偵斾傋偰GC娷検偺擇師揑側嬒堦壔偑恑傫偱偄傞偲峫偊傜傟傑偡丏巹偨偪偼丄儅僀僋儘愼怓懱傪傕偮懠偺唳拵椶乮僿價椶乯偲儅僀僋儘愼怓懱傪傕偨側偄唳拵椶乮儚僯乯傪傕偪偄偰丄偝傜偵徻嵶側夝愅傪恑傔偰偄傑偡丅傑偨丄崱屻偼丄偙偺傛偆側僎僲儉偺儌僓僀僋峔憿傪嶻傒弌偟偨梫場偲偟偰丄妀撪偺愼怓懱僥儕僩儕乕偵拝栚偟丄僋儘儅僠儞偺妀僟僀僫儈僋僗偲妀宆恑壔偺娭學偵偮偄偰尋媶傪峴偭偰偄偔梊掕偱偡丅
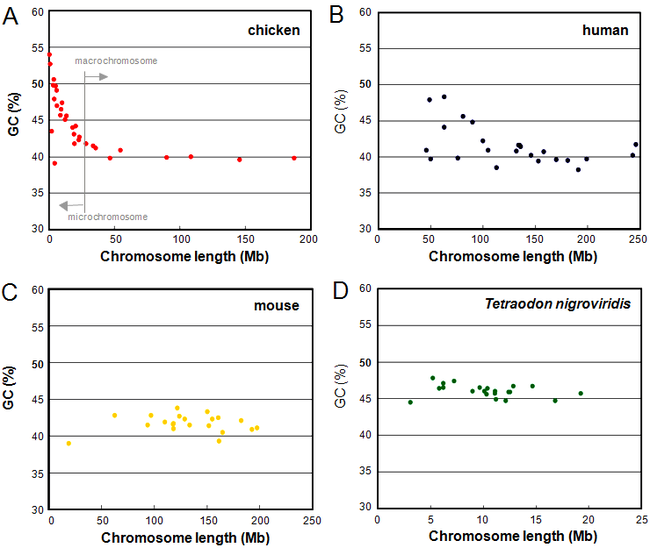
仾恾俉丂愼怓懱僒僀僘偲GC娷検偺娭學乮墶幉偑愼怓懱僒僀僘偛偲偺GC娷検丄廲幉偵奺愼怓懱DNA慡懱偺GC娷検傪帵偡丅乯
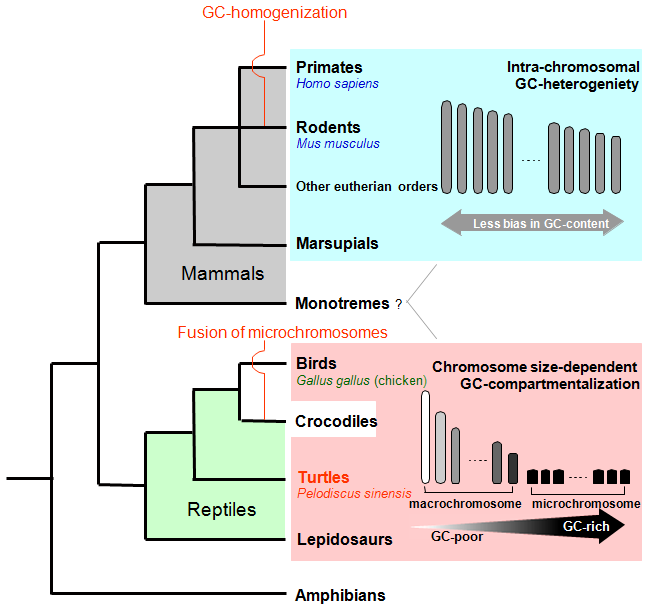
仾恾俋丂Chromosomal evolution and GC heterogeneity in amniote phylogeny