光合成と窒素固定の酸素パラドクス
シアノバクテリアの約半数は、空気中の窒素(N2)を窒素源として利用できる窒素固定能をもっています。窒素固定反応を担うニトロゲナーゼは、その特有の金属クラスターがO2に触れると秒から分単位で破壊されてしまうというO2に非常に脆弱な酵素です。このため、窒素固定能をもつ生物は、ニトロゲナーゼが作動する細胞内を嫌気的環境に保つために多様なメカニズムを進化させてきました。ところが、シアノバクテリアは、O2を発生する光合成で生きています。ということは、シアノバクテリアは環境のO2だけでなく、光合成で自ら作り出すO2に対しても防御する必要があるわけです。この光合成と窒素固定の“酸素パラドクス”を解決するために、シアノバクテリアは、他の生物とは異なる何か特別なメカニズムをもっているのに違いありません。
最も有名なメカニズムは、ヘテロシスト(heterocysts 異型細胞)と呼ばれる特別な細胞を分化するというもので、アナベナ(Anabaena)がその代表例です。しかし、ヘテロシストを分化せずに窒素固定を行うことができる、ヘテロシスト非形成型シアノバクテリア(nonheterocystous cyanobacteria)も多数存在し、海洋を含む多様な環境において生態系への窒素供給に重要な役割を担っています。Leptolyngbya boryana(レプトリンビア ボリアナ)もそのようなシアノバクテリアの一種です。ヘテロシスト分化と窒素固定は密接に連携しているため、これまでの研究の多くはヘテロシスト分化の分子機構に焦点が当てられていました。そのため、ヘテロシスト分化に関する転写制御タンパク質は同定されていましたが、窒素固定遺伝子群の発現を直接制御する転写制御タンパク質が未同定のまま残されていました。私たちは、窒素固定の制御に焦点を当てて研究を展開するために、L. boryanaを材料とすることにしました。L. boryanaは、藤田が1992年に形質転換系を確立しており(Fujita et al. 1992)、ヘテロシストを分化しないシアノバクテリアでは、ほぼ唯一分子生物学的な研究展開が可能な種になっています。

Anabaena sp. PCC 7120は、窒素枯渇に応答して窒素固定に特化した細胞ヘテロシスト(赤い矢印)を分化することができます。一方、L. boryanaは、窒素固定条件でも細胞は均一なままです。同じ細胞で光合成と窒素固定を両立させているようです。
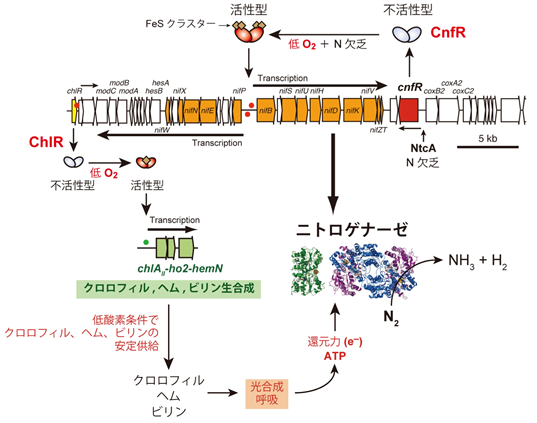
CnfRが窒素枯渇と低酸素に応答して活性化型に変換され、nifBとnifPの遺伝子間領域のプロモーターから両方向への転写を誘導します。一方、nif遺伝子クラスターには、低酸素でのクロロフィル生合成を可能とする3つの遺伝子を誘導するChlRをコードする遺伝子も含まれています。
私たちは、L. boryanaのゲノムに窒素固定に関連する遺伝子群が集中している窒素固定(nif)遺伝子クラスターを見つけました。この遺伝子クラスターに窒素固定遺伝子群の発現を制御するマスターレギュレータ遺伝子cnfR(cyanobacterial nitrogen fixation regulator)が存在することを見出しました (Tsujimoto et al. 2014)。遺伝子破壊株の形質などから、CnfRは低酸素を感知して窒素固定遺伝子群の転写を誘導する転写活性化タンパク質としてはたらくことがわかりました。また、この遺伝子クラスターのもう一方の端には、先にSynechocystis sp. PCC 6803で同定された低酸素条件下でもクロロフィル供給が可能となるように別の転写制御タンパク質ChlRがコードされていることも明らかになりました。窒素固定条件では、細胞内が低酸素から嫌気条件となることが必要とされます。そのような条件でクロロフィル生合成が滞ってしまうと、大量の還元力とATPを消費するニトロゲナーゼを駆動し続けることができません。ChlRによって低酸素でのクロロフィル・ヘム・ビリン色素の供給が可能となることで、ニトロゲナーゼの活性維持のため光合成と呼吸を稼働させることができるわけです。酸素パラドクスの統御という局面で、クロロフィルの低酸素適応機構と窒素固定の制御機構が収斂したことは、予想外の驚きでした。この成果は、名古屋大学ホームページ(NU Research)で紹介されました。
これらnif遺伝子群の転写誘導に際して、CnfRが認識するcis上流配列を同定するために、非窒素固定性シアノバクテリアSynechocystis sp. PCC 6803を活用したレポーター系を確立しました。この系を用いて、CnfRが認識するcis上流配列が、nifB上流、nifP上流およびorf118上流に存在することを明らかにしました(図3.1の赤いドットがそれらのcis上流配列を示します;Tsujimoto et al. 2016)。
文献
Fujita, Y., Takahashi, Y., Chuganji, M. and Matsubara, H. (1992) The nifH-like (frxC) gene is involved in the biosynthesis of chlorophyll in the filamentous cyanobacterium Plectonema boryanum. Plant Cell Physiol. 33, 81-92.
Tsujimoto, R., Kamiya, N., and Fujita, Y. (2014) Transcriptional regulators ChlR and CnfR are essential for diazotrophic growth of nonheterocystous cyanobacteria. Proc. Natl. Acad. Sci. USA 111, 6762-6767.
Tsujimoto, R., Kamiya, N., and Fujita, Y. (2016) Identification of a cis-element in nitrogen fixation genes recognized by CnfR in the nonheterocystous nitrogen-fixing cyanobacterium Leptolyngbya boryana. Mol. Microbiol. 101, 411–424.
藤田祐一、辻本良真 (2016) 酸素パラドクスの統御で収斂するクロロフィル生合成と窒素固定 光合成研究 26, 204-215.