Laboratory of Animal Genetics and Breeding

非コード領域に存在する遺伝子発現制御機能の解明とその制御法の研究による有用動物の開発
・脊椎動物ゲノムに特有のdistal enhancerによる遺伝子発現調節機構の研究 ( PNAS 2002, 2003, 2008, Development 2009, Nature 2012, etc.)哺乳類の場合、ゲノム中に遺伝子が占める割合は数%以下で、90%以上の領域は遺伝子をコードしていないためかつては「ジャンク」と呼ばれたこともありましたが、今では遺伝子発現の制御を始めとする重要な機能を担っていることが次々と明らかになってきています。 まだその多くは機能未知のまま残されており、手つかずの「宝の山」と言って過言ではありません。有用動物を開発するにあたって、タンパク質をコードする領域のゲノム操作では実現できない表現型の操作が可能になると考えられます。
脊椎動物ゲノムの特長として、ゲノム非コード領域に存在するdistal enhancer の存在が挙げられます。プロモーター周辺のcis-elementだけではなく、場合によっては数百kb以上も離れた場所に存在するエンハンサーエレメントによって精緻な制御が行われています。 近年、GWASによる疾患関連SNPの多く(95%以上と言われています)がこのようなdistal enhancer に存在することが明らかになり、その遺伝子発現調節機構における重要性が一般的に認識されるようになりました。 このような長距離の転写制御はCTCFなどのファクターによるループ形成や、自己形成するTAD (Topologically Associating Domain) 構造に依存していると考えられています。 当研究室ではこれまでにBACを用いたdistal enhancerの機能解析とその性質・進化について研究を行ってきました。さらに最近ゲノム編集技術の進展によりTAD構造に直接改変を加えて機能解析ができるようになっています。 TAD内のdistal enhancerの働きを理解し応用することで、発現調節機能を変更する方法を開発し、有用動物開発に役立てることを目標に研究を進めています。
また、distal enhancer の進化的変更が動物進化に本質的な役割を果たすことが現在注目を集めています。 私たちは特にdistal enhancerの進化についてトランスジェニックマウスを使用する解析法について先駆けとなる研究を行い、これまでに国内外の多くの共同研究も含め分野を牽引する成果を上げてきています(哺乳類トランスポゾン由来エンハンサーの発見と実証:東工大岡田典弘研究室(当時)との共同研究、など)。 トランスポゾン挿入や遺伝子変換などによる新しいcis-element成立の条件を知ることにより、進化に寄与するcis-elementがどのように生じ、消えてゆくのか知ろうとしています。長大な非コード領域は脊椎動物の特徴的な進化の産物ですが、ただの無意味な配列ではなく積極的に新機能を取り込むために進化した結果の「進化の仕組み」であると考えて研究に取り組んでいます。 実験による実証アプローチにより cis-element 駆動型の進化機構を解明していきます。このような進化機構を明らかにし応用することで、有用動物のこれまでにない開発方法に結びつけようとしています。
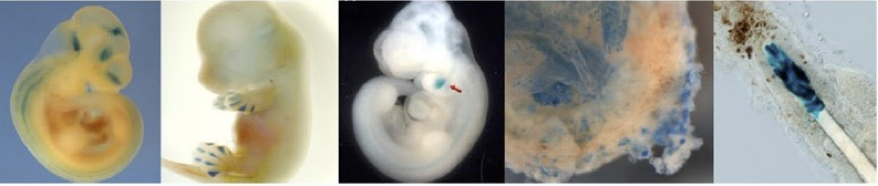
図1 エンハンサー活性の in vivo レポーターアッセイによる解析
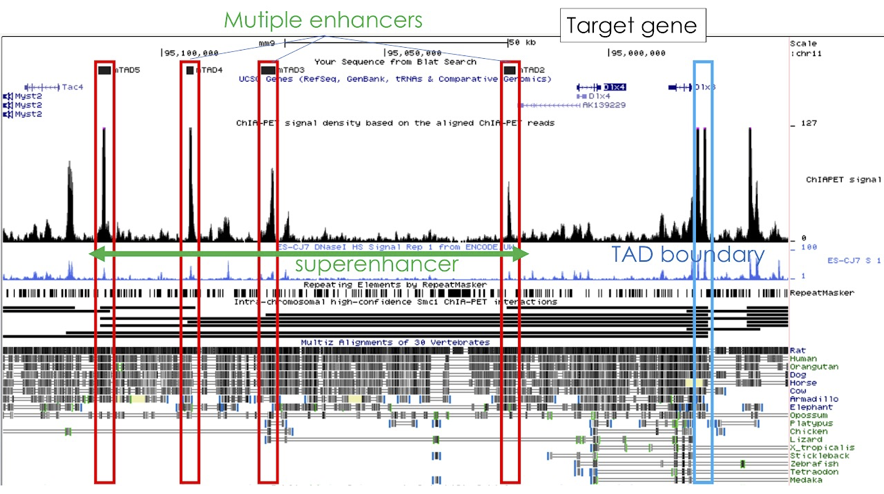
図2 遺伝子発現を制御しているゲノムの非コード領域の構成例
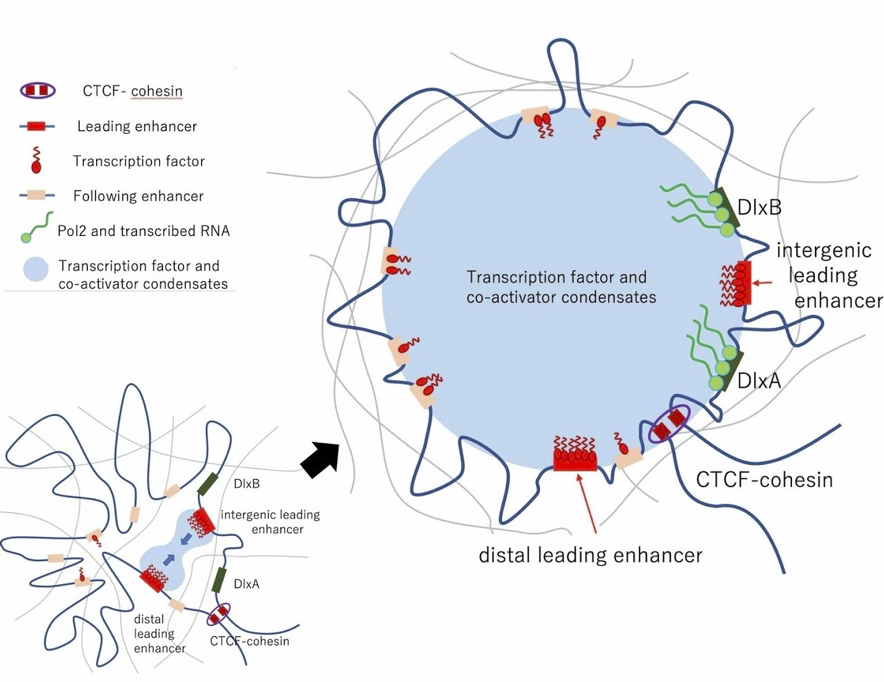
図3 スーパーエンハンサーの実態としてのMolecular condensate のモデル図。リーディングエンハンサーは最初にbiomolecular condensate の「核」を作る役目をしており、これが融合して巨大な biomolecular condensate に成長すると考えられる。 このMolecular condensate 形成の制御が遺伝子発現と動物表現型操作のための重要なターゲットになる。 参照:Kenta Sumiyama, Akira Tanave. "The regulatory landscape of the Dlx gene system in branchial arches: shared characteristics among Dlx bigene clusters and evolution." Development, growth & differentiation (2020) 62(5): 355-362. DOI: 10.1111/dgd.12671