|
 |
|
|
|
 |
 |
 |
 |
 |
 |
 |
|
セスキテルペンとはイソプレン単位が3つ重合してできる骨格をもつ化合物のグループで、医薬品、香料、毒素などの二次代謝産物が含まれます。穀類に感染する糸状菌フザリウムは、セスキテルペン毒素であるトリコテセン系かび毒を穀粒中に蓄積させ、食の安全を脅かします。重金属や農薬のような他の食の危害要因とは異なり、その側鎖構造は菌とともに進化し、新しいトリコテセン系かび毒が突如として出現することが知られています。私たちのグループではトリコテセン生合成遺伝子の機能解析を通じ、その側鎖多様性を生み出す進化機構を研究しています。また、複雑な外部環境要因に応答して、大量にセスキテルペン毒素を生産することのできるフザリウムの遺伝子発現制御メカニズムを転写因子の機能の点から調べています。 一方、糸状菌には二次代謝産物を通して私たちの生活に関わるだけでなく、付加価値の高い酵素や有機酸を大量に生産し、発酵食品の製造に用いられる有用な菌が存在します。その代表的な微生物が酒、味噌、醤油の製造に使われている麹菌やその仲間で、農産物を栄養として旺盛に生育するために、細胞壁やタンパク質を効率的に分解します。これらの
Aspergillus 属のカビは古くから飲料、食品の製造に使われてきました。私たちのグループでは、これまで利用されてきたデンプン分解やタンパク質分解以外に、ゲノム情報から予想される新たな機能の制御・利活用を通じ、食・環境に関わる諸問題の解決に貢献する研究を進めています。
(1)トリコテセン系毒素の側鎖多様性を生み出す遺伝学的メカニズムの解明  図1:トリコテセン骨格と側鎖。フザリウムでは C-3 位が必ず(2)または(3)となる。(I)T-2 toxin、(II)4,15-diacetylnivalenol、(III)NX-3 トリコテセンは比較的単純な基本骨格(図1)を持つセスキテルペンで、穀粒中に蓄積し、食の安全を脅かします。その骨格の5カ所の炭素のうち、どの位置にどのような置換基のついたトリコテセンをフザリウムが最終産物として生産するかは遺伝的に決まっています。これは生合成酵素の性質や機能が、祖先種からより多様化するように進化して来たためです。水酸基、アセチル基の単純な置換基だけでも組み合わせパターンは理論上 4 x 2 x 3 x 3 x 2 = 144通り(図1)ですが、既知のフザリウムが生産するトリコテセン側鎖のパターンは実際には限られます。 (I)、(II)に示す2つのトリコテセンは異なるフザリウム種が生産し、そのC環の構造は全く同じです 私たちの研究グループでは生合成シャント化合物を多様なフザリウムの生合成遺伝子と置換した株へフィーディングし、代謝産物の解析を行っています。側鎖修飾に関わる酵素の性状を解析し、生合成経路の精密解析を進めることによって、トリコテセン側鎖多様性を生み出す遺伝学的メカニズムを明らかにするとともに、新たな食の危害要因の低減化に役立てるための研究を行っています。 (2)セスキテルペン産生制御機構の解析 フザリウムは pH、糖、水分活性、窒素源などの環境要因を感知し、トリコテセン生合成遺伝子(Tri 遺伝子)の発現を制御します。Tri 遺伝子の大部分は遺伝子クラスター(Tri 遺伝子クラスター)を形成していますが、そのコア領域に存在する Tri6 遺伝子は、Cys2His2 型の3つの zinc finger をもつ正の転写因子をコードするトリコテセン生合成に必須の遺伝子です。Tri6p転写因子は、Tri遺伝子のプロモーター領域(Tri6自身を含む)に存在するコンセンサス配列 YNAGGCC へ結合してTri 遺伝子の転写を活性化しますが、そのためにはまずTri6 自身が Tri6p 転写因子に依存しないで転写(Tri6p-independent initial transcription: Tri6p 非依存性初期転写)され、翻訳産物 Tri6p として細胞内に存在する必要があります。さらに Tri6p 転写因子による Tri6 の転写活性化(Tri6p-dependent positive feedback
activation: Tri6p 依存性自己転写活性化)には、これまでの研究からタンパク質翻訳後に Tri10p を含む何らか因子の作用を受けて活性化される必要があると考えています。私たちのグループでは、Tri6p 非依存性初期転写と Tri6p 依存性自己転写活性化を区別できる系を構築し、どのような外部環境要因がこれらの異なる様式の転写を制御しているのかについて研究を進めています。
また、フザリウムの生育に影響を与えずに、Tri6 の転写のみに影響を与える化合物の探索を行い、Tri6 発現を促進もしくは抑制する化合物をいくつか見出しています。これらの化合物は、毒素生産の抑制剤もしくは有用セスキテルペンの生産促進剤としての応用が期待されますが、その作用点の探索を進めることによってTri6発現活性化調節の分子機構を明らかにし、セスキテルペンの発現制御機構を巧みに利用した異種セスキテルペンの代謝工学へと発展させることを目指しています。 (3)フザリウムの特定の遺伝子の変異株ライブラリーの作成系の構築 ゲノムサイエンスやインフォマティックスの著しい進展に伴って、これまで多くの遺伝子の機能が解析されてきました。しかし、特定の遺伝子に対象を絞ってアミノ酸置換変異を導入し、興味のある表現系を示す変異株をスクリーニングすることができるツールはありません。フザリウムのセスキテルペン生産制御において鍵となる転写因子をコードする Tri6 やヘミセルロースの分解酵素遺伝子の発現制御に関わる転写因子をコードする遺伝子などの特定の遺伝子にターゲットを絞り、細胞内で直接、ランダムに遺伝子改変を施した変異株ライブラリーからより好ましい形質を示す変異遺伝子を人工的に進化させて選抜できるシステムの開発を目指して研究を進めています。 将来的には食品・農業廃資源などを原料に、毒素の代わりに有機化学合成ではコストがかかるセスキテルペン医薬品原料などを大量に生産させる宿主としたフザリウムの代謝工学の基盤形成を目指して研究を進めています。
(4)コーヒー残渣 SCG(spent
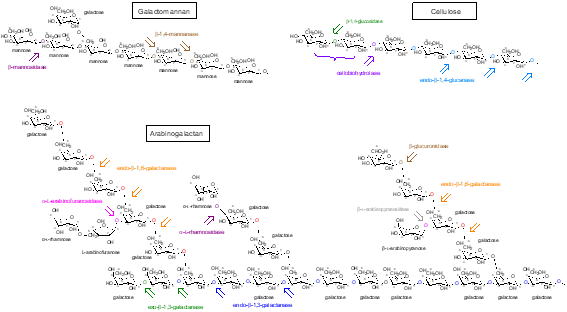
coffee ground)の効率的分解を目指した基盤研究および SCG を利用した発酵食への応用に関する研究 コーヒーは石油に次いで第2位の国際貿易額を誇る、という怪しげな都市伝説が出回るほど親しまれている身近な飲料で、世界中で広く消費されています。嗜好品としての側面のみならず健康増進に役立つ様々な機能性成分を含み、血中コレステロールや血糖を低下させる効果、腸内善玉菌(ビフィズス菌)の増殖を賦活化する活性、免疫機能の向上など様々な可能性が報告されています。 コーヒーを熱水抽出した後の残渣は SCG(Spent Coffee Ground)と称され、これまで大部分が廃棄されてきました。21世紀に入り、廃棄物削減、資源循環、温暖化対策など、地球規模で対策を講ずべき様々な環境問題へ対処するため、企業も近年は様々な取り組みを行っています。SCG は元来、多糖、リグニン、タンパク質、ミネラルなどを豊富に含むことから、優れた活用法の開拓が期待されますが、未だ循環資源として十分に有効活用する方法の開発には至っていません。この理由としては、不溶性で難分解性の SCG を断片化し、利用可能なオリゴ糖、糖タンパク質、ポリフェノールへと変換する手段が欠如していることが挙げられます。 SCG の糖の主成分は、セルロース、ガラクトマンナン、Ⅱ型アラビノガラクタン(図2)で、一般に水分活性が低い条件下でも生育することのできるカビはこれらの多糖を断片化する酵素を菌体外へ分泌し、栄養分として利用します。そこで糸状菌の中でも生育が早くモデル微生物としても研究が進んでいる麹菌 Aspergillus oryzae を用い、SCG を栄養として利用して菌体バイオマスを増やす上で役立つ遺伝子の機能やII型アラビノガラクタンの分解制御機構について調べています。この研究を通じ、SCG のアップサイクリングに必要な効率的糖化の学術的基盤の構築を目指します。また基礎研究だけではなく、実際に SCG を混ぜたテンペやパンなどの発酵食を作成し、味覚や機能性の評価を行って研究室員で楽しんでいます。
図2:SCG を構成する糖の構造 |
|
Copyright © 2016 応用微生物学研究分野 All Rights Reserved. |