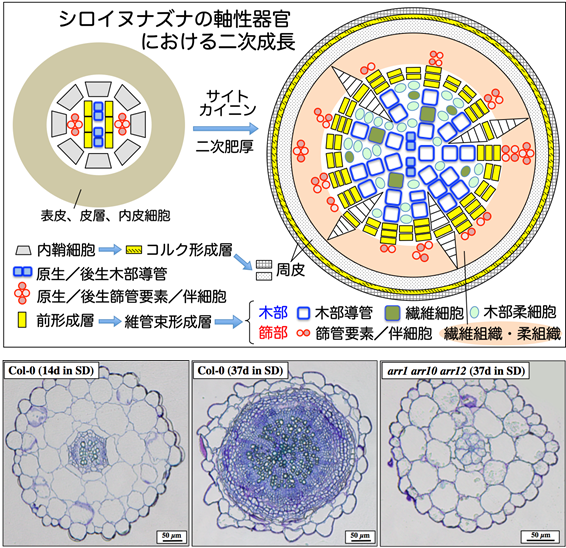
種子植物の二次成長
植物は生涯を通じて根、茎、葉、花などの器官を発生させ続けながら成長します。多くの種子植物の体づくりは、器官原基の形成とその発生を司る一次成長と、発生した器官の体軸肥厚を司る二次成長とに分けられます。一次成長は頂端分裂組織に由来する細胞が分裂・分化を経て器官形成しながら主に基部−先端軸方向へ伸長する成長様式です。一方、単子葉を除く種子植物の主に根、胚軸などの軸性器官で観察される二次成長は、これらの器官の中に存在する前形成層と内鞘細胞が分裂活性を獲得して体軸が中央-周辺軸方向へ肥厚する成長様式です。モデル高等植物シロイヌナズナでは草本植物に典型的な二次成長が観察されることが知られています。二次成長が始まると前形成層は木部・篩部(維管束系)に分化する細胞を生じさせる維管束形成層とよばれる分裂組織として、内鞘細胞は自分の細胞の数を増やしながら、コルク形成層とよばれる周皮を作る細胞を生成する幹細胞として機能するようになります。その結果、成熟した植物組織の大部分は維管束形成層とコルク形成層における細胞分裂に由来する軸性器官の細胞よって占められます(図参照)。すなわち、二次成長は器官内および器官間の高次の維管束系ネットワークを構築しつつ、一次成長により発生した新規の器官を維持するための体軸強度を担保するために必須の役割を果たしています。したがって、二次成長における細胞分裂活性調節と分化運命決定機構は、植物の体制構築における二次組織の肥厚、並びにその構造制御に密接に関連していると考えられます。
当研究室では、シロイヌナズナの二成分制御系のゲノムワイド解析に着手し、植物ホルモンとして有名なサイトカイニンのシグナルはHis Kinase受容体→HPt仲介因子→Response RegulatorへのHis-Aspリン酸リレーを介して伝達されることを明らかにしてきました。この過程で、サイトカイニン情報伝達系が欠損(arr1 arr10 arr12三重変異体)すると本来分裂活性が備わるべき前形成層と内鞘細胞の細胞分裂が活性化されないために二次成長が誘導されないことを見いだしました(図参照)。サイトカイニンは細胞分裂の活性化を誘起する植物ホルモンとして古くから知られています。私たちはサイトカイニンシグナルの下流で発現する如何なる機構が背景となって細胞分裂活性が維持されるのかというサイトカイニンの生理作用に関する最も本質的な疑問を明らかにしたいと考えています。植物の二次成長(肥厚)制御は農業上の重要形質とされる作物の直立性向上や植物バイオマスの高資源化を支えるグリーンイノベーション技術に直結すると期待されます。