植物時計のしくみとはたらき
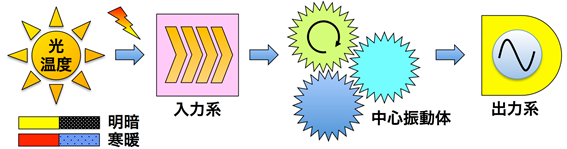
動物、植物、菌類、藻類(以上、真核生物)、光合成細菌(原核生物)を含むほとんどすべての生物には概日リズム(circadian rhythm)とよばれる約24時間周期で変動する生理現象が観察されます。概日リズムは生物が昼夜の交代や季節の移ろいという地球環境の周期的変化に適応するうえで、優位性をもたらす生命機能であると考えられています。概日リズムの種類は生物種によって多様ですが(例えば、哺乳動物では体温、睡眠覚醒、ホルモンの分泌、摂食など、高等植物では胚軸の伸長、葉の就眠運動、気孔開閉、光合成などが知られています。)、時間を計測するための機構として非常に重要な以下の普遍的性質を備えています。概日リズムは外部環境の変化が無くても約24時間周期のリズムが自律的に継続します。その周期は生理学的な温度範囲において、通常の化学反応と異なり、温度変化の影響をあまり受けません。光および温度による外部からの刺激などに対してリズムの位相を変化させることで昼夜の変化に同調することができます。このような特性を保持する概日リズムを生成するための発振装置としての役割を果たしているのが概日時計であり、そのなかで中心振動体として機能するタンパク質の活性および存在量の周期的変化が生物時計の振動に相当することが知られています。概日時計には中心振動体に光と温度などの外部情報を伝達し、時計の振動位相を調節するはたらきを担っている入力機構や中心振動体からの時間情報に応じて決まった時間に決まった応答を誘導あるいは抑制するための出力機構が存在することが知られています。本研究室では、高等植物シロイヌナズナを対象に植物時計の分子機構を研究しています。
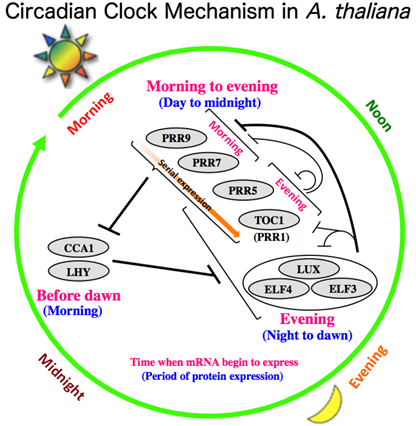
シロイヌナズナの概日時計の中心振動体は3つのクラスの転写制御因子CCA1/LHY, PRR9/7/5/TOC1, ELF4-ELF3-LUXから構成されていると考えられています。中心振動体をコードする遺伝子発現に関して、CCA1/LHYは夜明け前に、PRR9/7/5/TOC1は朝から夕方にかけて、ELF4/ELF3/LUXは夕方から夜にかけてmRNAが一過的に誘導されることが知られています。mRNAを鋳型に翻訳された中心振動体タンパク質は転写抑制因子として機能し、自分の発現位相よりも前に発現している中心振動体遺伝子の発現を負に制御します。その結果、自分の発現位相よりも後に発現する遺伝子の抑制が解除されます。これが繰り返されることにより、CCA1/LHY→ PRR9/7/5/TOC1→ELF4-ELF3-LUXの順序で1日の時間経過とともに中心振動体遺伝子が時間特異的な発現ピークをもって振動するというモデルを概日時計の基本機構として提唱しています。これを基盤に概日時計の特性である自由継続性、温度補償性、同調性を分子レベルで理解することを研究の目的としています。
概日時計は1日の時間を計測し、決まった時間に決まった応答を誘導することにより、1日周期の生物リズムを生成するための機構であるだけでなく、季節という1年周期の日長や温度サイクルに対する応答をも支配しています。シロイヌナズナは長日植物に属し、実験室で明期の時間帯設定を変えることにより、花成を制御することが可能です。野生型のシロイヌナズナは栽培条件を整えると長日条件では3週間から4週間で栄養成長から生殖成長へと成長相が転換しますが、短日条件では顕著に花成が遅延します。時計機構が異常になった変異体では長日条件における花成が遅延したり、その逆に、花成が誘導されない短日条件においても、花成が促進されたりすることが知られています。概日時計の振動はどのような機構で出力されて多様な生物リズムの生成に結びついているのかを理解することは植物の能力を活かす上で重要な研究課題であるといえます。
