犬飼グループ
現在の研究テーマ
(1) 天水田での乾燥回避に重要な根系形態の把握 |
(2) イネ根系の分子メカニズムの解明 |
(2-5) イネのembryonic root(幼根)とpost-embryonic roots(冠根および側根)の発生はそれぞれ異なる遺伝制御を受ける |
(1) 天水田での乾燥回避に重要な根系形態の把握
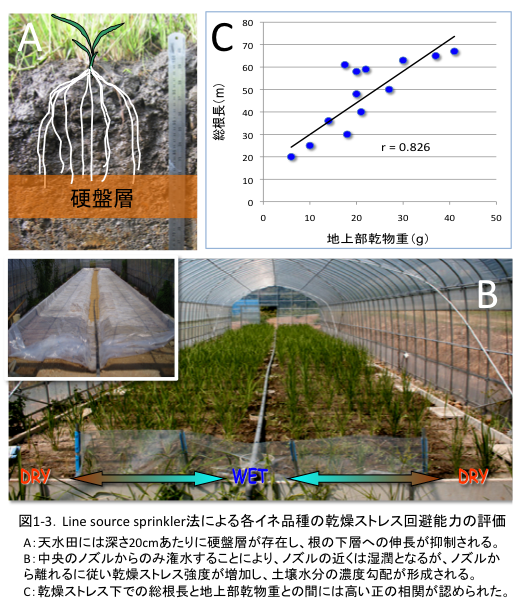
(1-1) 高い乾燥回避能力はストレス下での根長の維持・増加によりもたらされる
天水田には深さ20cmあたりに硬盤層が存在し、これにより根の下層への伸長が抑制されます(図1-3A)。硬盤層は不透水層であるため、これより下層が湿潤状態であっても根はその水を吸収することができません。このような土壌環境下において重要な根系形態を探るために、私たちは土壌深度20cm部位にビニールシートを敷き、根の下層への伸長と水の上層への移動を阻害できる圃場を設定しました(図1-3B)。この圃場において、湿潤から乾燥まで連続的な土壌水分勾配を発生させることを可能にするLine source sprinkler 法を導入し、様々なイネ品種を評価しました。その結果、ストレス下での総根長と地上部乾物重との間には有意な正の相関が認められたため(図1-3C)、高いストレス回避能力は乾燥ストレス下での根長の維持や増加によりもたらされると考えられました。
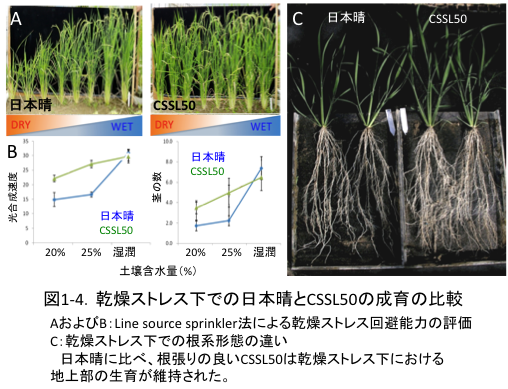
(1-2) ストレス下での根長の維持・増加は、限られた水の積極的な吸収を通して光合成量の減少を軽減する
一方、地上部形態の大きく異なる品種間において耐旱性を比べる上では、根系発達の違いが実際にどの程度ストレス回避に貢献しているのかを判断するのは困難です。そこで、お互いの遺伝的背景が非常に近い染色体断片置換系統群(CSSL、農業生物資源研究所イネゲノムリソースセンターより分譲)を用いてストレス回避能力を評価しました。その結果、乾燥ストレス下での地上部成育の維持能力が最も高い系統としてCSSL50が選抜されました。本系統の乾燥ストレス下における総根長は、反復親品種である日本晴の約2倍にまで達し、また光合成速度は乾燥下においても高く維持されていました。(図1-4)。以上より、乾燥ストレス下において根の長さを維持、あるいは増加できる能力が、限られた水の積極的な吸収を通して光合成量、ひいては乾物生産の減少を回避する上で非常に重要であることが確認されました。
(2) イネ根系の分子メカニズムの解明
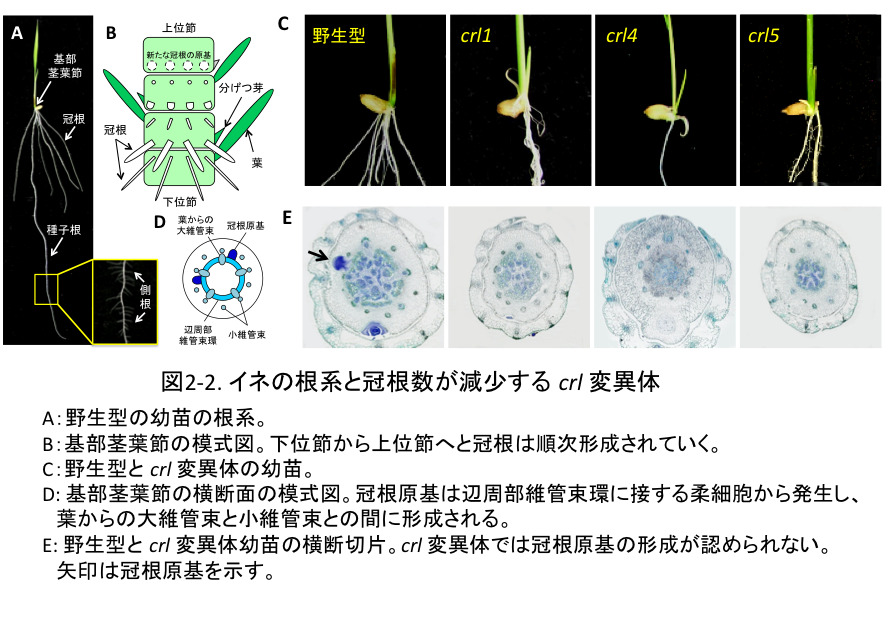
(2-1) イネの根系と、その形態に異常が見られる突然変異体
イネの根系は主根 (種子根)、不定根 (冠根) およびそれらから発生する側根によって構成されています(図2-2A)。胚発生の過程で種子根(幼根)が形成され、地上部の発育に伴い、基部茎葉節から冠根が次々と発生します (図2-2B)。私たちは、これら個々の根の発生に異常を示す突然変異体を材料に(図2-2C、E)、イネの根系がどのような遺伝的メカニズムによって形作られるのかを解析しています。今までに明らかになったことは、次のようなことです。
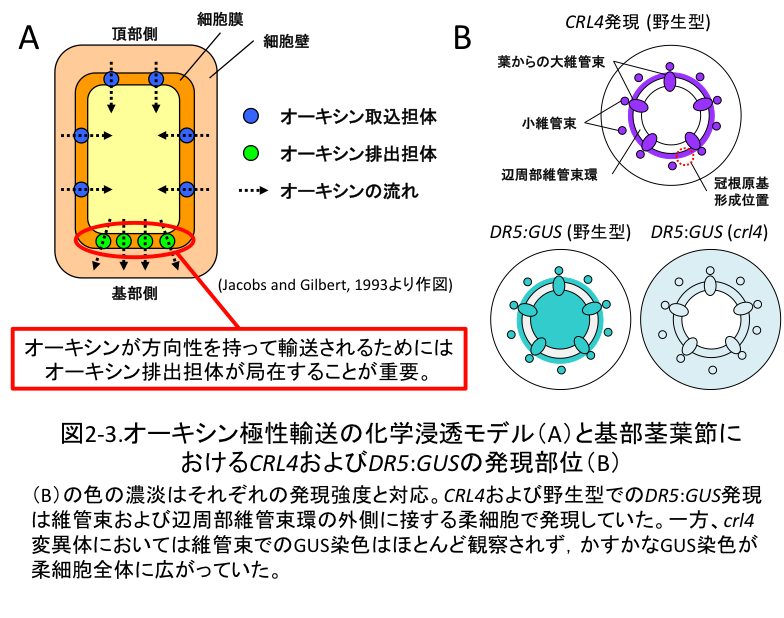
(2-2) CRL4遺伝子は適切なオーキシンの集積と濃度勾配の確立を通してオーキシン信号伝達を進行させる
crl4変異体の原因遺伝子を単離したところ、CRL4/OsGNOM遺伝子はオーキシン極性輸送に関与するArf-GEFをコードすることが分かりました。GNOM/EMB30は、オーキシン排出キャリアーであるPIN1のリサイクルを制御していることが報告されています(図2-3A)。そこでcrl4変異体におけるオーキシン輸送能をトレーサー実験により解析したところ、crl4変異体では野生型に比べ著しく輸送能が低下していました。また、植物体内でのオーキシンの局在をDR5:GUSにより可視化したところ、野生型では維管束と辺周部維管束環に接する柔細胞でGUS染色が観察され、この発現パターンはin situ hybridizationにより解析したCRL4/OsGNOMの発現パターンと非常によく一致していました。一方、crl4変異体においては維管束でのDR5:GUS染色はほとんど観察されず、微かなGUS染色が柔細胞全体に広がっていました (図2-3B)。
以上より、CRL4遺伝子によるオーキシン濃度勾配の形成が根の発生には必須であることが明らかになりました。
(2-3) CRL1遺伝子はオーキシン信号伝達経路下で機能する
オーキシン信号の伝達は、 AUX/ IAAタンパク質とARFタンパク質の相互作用によって制御されていることが報告されています(図2-4)。オーキシンが存在しないときはAUX/ IAAとARFがヘテロダイマーを形成し、ARFの転写活性能が抑制されてオーキシン信号は伝達されません。一方、オーキシン存在下ではAUX/ IAAが分解されます。その結果、ARFの転写活性能が回復してオーキシン信号が下流へと伝達され、最終的に根の発生が誘導されます。しかし、オーキシン信号伝達と根の発生との間を結び付ける分子メカニズムはミッシングリンクとして残されていました。
私たちはcrl1変異体の原因遺伝子の単離を試みた結果、CRL1/ARL1 はAS2/LOBタンパク質をコードし、またオーキシンにより発現が誘導されることが分かりました。そこで、本遺伝子とAUX/ IAAおよびARFタンパク質によるオーキシン信号伝達経路との関係を詳細に解析した結果、CRL1/ARL1はARFの直接的な標的遺伝子であることが判明しました (図2-4)。同様に、シロイヌナズナの側根形成においても、CRL1/ARL1と相同性の高いAS2/LOBタンパク質であるLBD16およびLBD29がARF7およびARF19の直下に位置し、最終的に側根形成を促すことが報告されました (図2-4)。また、crl1変異体でも側根数の減少が認められるため、オーキシン信号伝達と冠根および側根の発生を結び付ける正の制御因子として、AS2/LOBタンパク質が機能していることが明らかになりました。
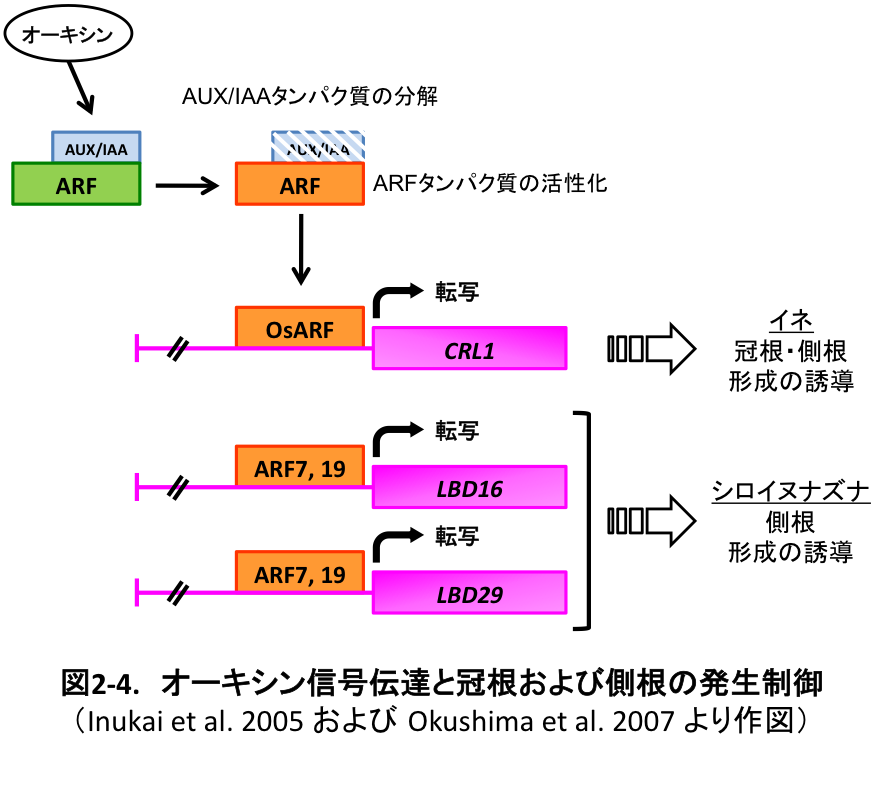
(2-4) オーキシンにより発現誘導を受けたCRL5遺伝子は、サイトカイニン信号伝達を負に制御する
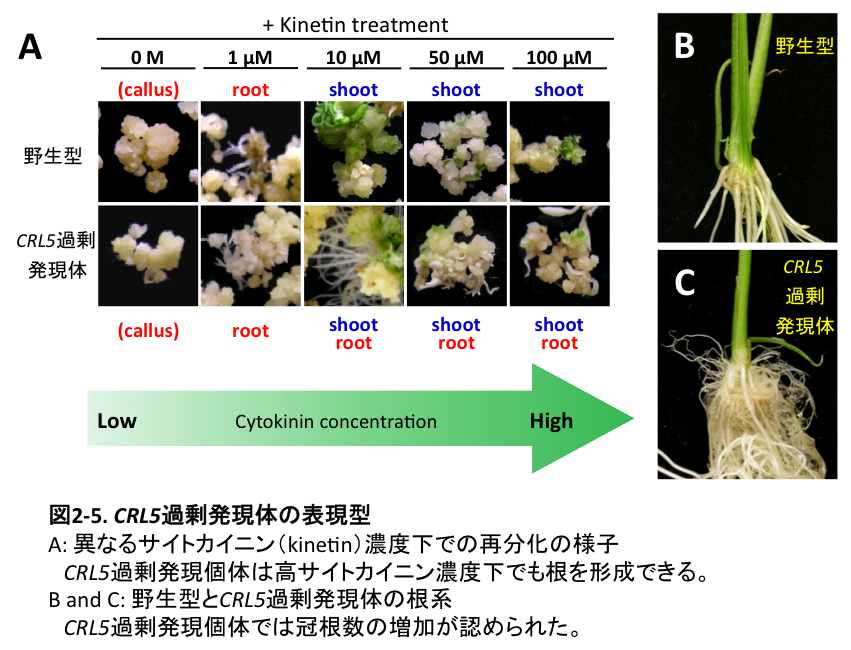
地上部(シュート)諸器官や根の発生には、植物ホルモンであるオーキシンとサイトカイニンが重要な役割を果たし、両者は拮抗的に作用することが古くから報告されています。例えば、カルスからの器官の再分化時において、サイトカイニン濃度が相対的に高ければシュートが、逆にオーキシン濃度が高ければ根が形成されます(図2-5A上段)。しかし、これら両ホルモン間の拮抗作用を支配する分子機構の実体は未解明の点が多く残されていました。
私たちは、crl5変異体の原因遺伝子の単離を試みた結果、CRL5はAP2型転写因子をコードすることが分かりました。本遺伝子の発現はオーキシンにより誘導され、またCRL1/ARL1遺伝子と同様にARFの直接の標的因子であることが示唆されました。このCRL5遺伝子の過剰発現個体は根の発生数が増加し、かつサイトカイニンによる根形成の抑制作用に対して耐性を示しました(図2-5A)。
そこで、サイトカイニン信号伝達に関与する遺伝子群の発現解析を行った結果、負の制御因子であるOsRR1遺伝子などのType-A Response Regulatorの発現が野生型に比べcrl5変異体で低く、逆にCRL5遺伝子の過剰発現体で高い傾向が認められました(図2-6A)。さらに、crl5変異体においてCRL5プロモーター下でOsRR1を発現させた形質転換体では、crl5変異体の冠根数の減少が回復することが分かりました(図2-6B)。以上より、オーキシンにより誘導を受けたCRL5遺伝子は、サイトカイニン信号伝達を負に制御することを通して冠根の発生を促すと考えられ、両ホルモン間の拮抗作用機構において本遺伝子が中心的な役割を果たしていることが判明しました(図2-6C)。
また、前述のように根の発生にはAS2/LOBタンパク質をコードする遺伝子群が重要なことが明らかになっています。しかし、これらの遺伝子群の過剰発現体では根の発生数の増加が認められず、品種育成に利用可能な有用遺伝子の発見が切望されていました。これに関して、CRL5過剰発現体では顕著に冠根数が増加したため (図2-5B、 C)、今後、根系形態の人為的改良に向けた本遺伝子の有用利用が大いに期待されます。

(2-5) イネのembryonic root(幼根)とpost-embryonic roots(冠根および側根)の発生はそれぞれ異なる遺伝制御を受ける
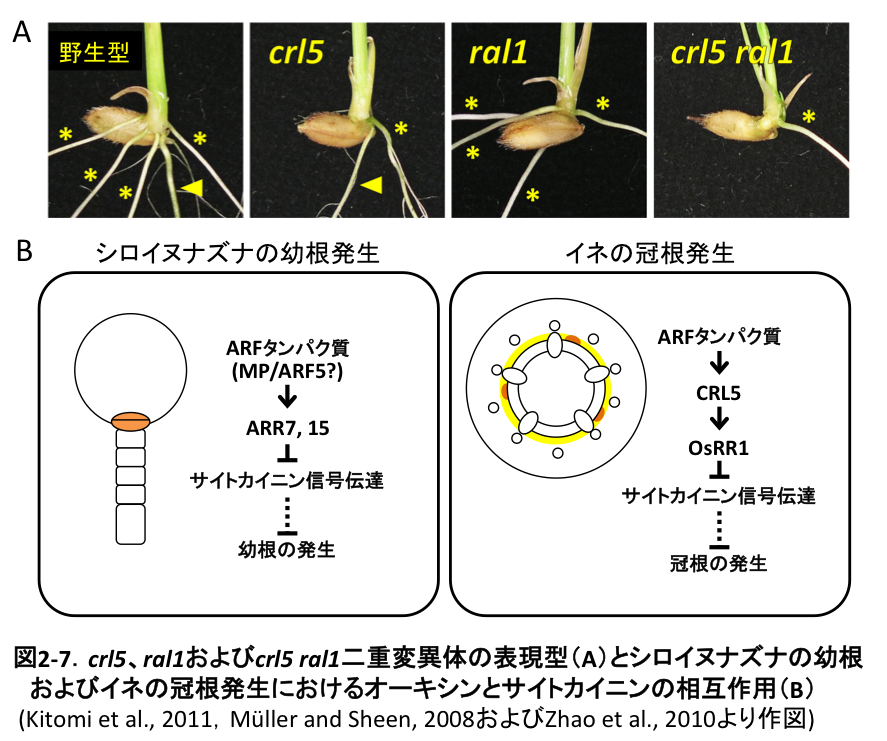
根は大まかに、胚発生の段階で形成されるembryonic rootと胚発生後に発生するpost-embryonic rootの2種類に分類できます。イネにおいて冠根および側根はpost-embryonic root、幼根はembryonic rootに分けられ、幼根のみが欠損する突然変異体としてradicleless1 (ral1) 等が報告されています (図2-7A)。一方、前述のcrl変異体では冠根や側根の発生に異常が生じるにも関わらず、幼根は正常に発生していました (図2-7A)。また、crl5 ral1二重変異体では幼根 (図2-7A、矢頭) が欠失し、冠根 (図2-7A、アスタリスク) はcrl5変異体と同程度に減少するという相加的な表現型を示しました。以上より、イネにおいてembryonic rootの発生とpost-embryonic rootの発生にはそれぞれ異なった制御機構が存在すると考えられます。上述のように、イネの冠根発生ではOsARFとOsRR1との間でCRL5が機能しています。一方、シロイヌナズナの幼根発生においてはMONOPTEROS (MP)/ARF5が直接ARR7およびARR15の発現を正に制御すると考えられています (図2-7B)。CRL5過剰発現体では、冠根数は増加するものの幼根数の増加は認められなかったことから、イネの幼根発生においてもOsARFとOsRRが直接相互作用していると推定されます。
幼根は、特に成育初期の養水分の吸収や植物体の支持を一手に担う非常に重要な器官です。今後、この幼根の発生のしくみを明らかにできれば、冠根や側根などのpost-embryonic rootsと併せた根系形成機構の包括的な理解が飛躍的に進むとともに、幼根数の人為的制御を可能にする有用遺伝子の同定へと繋がるものと期待されます。
![]()