研究内容Research subjects
つぼみを開花させるしくみの研究
花咲か爺さんの灰を科学する
寒い冬を堅い冬芽の中で過ごしたサクラのつぼみは、春になるとふくらみ始め、やがて開花を迎えます。このとき、休眠しているつぼみを目覚めさせ、開花に向かわせる「開花ホルモン」が働いていると予想されます。昔話の「花咲か爺さん」がまいた灰のような働きを持つ物質を植物自身が作るのではないかということです。でも、まだその実体はよくわかっていません。
ほかの植物ではどうでしょうか、シロイヌナズナのつぼみは連続的に大きくなり、休眠することなく開花します。しかし、ジャスモン酸という植物ホルモンを作れない突然変異体では、つぼみの成長が開花直前で停止し、咲かなくなってしまう(または開花が遅れる)ことがわかりました。この植物にジャスモン酸をかけると開花するようになります。同様の現象がナタネ、タバコ、イネなどでも見つかっており、ジャスモン酸は多くの植物で「開花ホルモン」として働いていることがわかってきました。ただしオーキシンやジベレリンも開花促進に関係すると言われていることから、実際の制御はもっと複雑かもしれません。
現在は、十分成長したつぼみではどのようなしくみでジャスモン酸の生合成が活性化されるのか、作られたジャスモン酸がどのようにして花弁の伸長や雄しべ・雌しべの成熟を実現するのかを調べ、開花という現象の遺伝子レベルでの理解を目指しています。一方、ジャスモン酸は植物が害虫から身を守る防御応答でも重要な働きを持っています。防御応答と開花制御で共通のしくみが働いているのかどうかについても調べています。


(図1) ジャスモン酸を作ることができないdad1 突然変異体のつぼみは開花しない(左)が、ジャスモン酸をかけると野生型と同じように開花する(右)。
花粉表層構造の形成機構の研究
植物のナノテクノロジー
電子顕微鏡で見た花粉は、網目状、突起状、流紋状など種ごとに異なる特徴的な姿をしています。この形を作っているのはエキシンと呼ばれる花粉の外殻構造で、スポロポレニンという名前の脂質由来のバイオプラスチックでできています。エキシンは虫や風に運んでもらうための「軽さ」と内部の雄性配偶子を保護する「頑丈さ」を兼ね備えた優れた構造物であると考えられます。
では、このナノスケールの立体構造を植物はどうやって作るのでしょうか?
私たちは正常なエキシンが作れなくなったシロイヌナズナの突然変異体を探しました。見つけた突然変異体にkaonashi1(kns1)、kns2、kns3、と名前をつけ、これまでに約40個のkns突然変異体を見つけています。変異を起こしていた遺伝子を調べてみると、その多くがペクチン、キシラン、アラビノガラクタンなどの多糖の生合成に関係する遺伝子でした。多糖がエキシンの立体構造を作るのに何らかの働きを持っているということになります。そのしくみはまだよくわかりませんが、多糖がエキシンを成形する鋳型のような働きをしているのではないかと考えています。では、この鋳型はどのようにして作られるのでしょうか。また、種ごとに異なるエキシンの構造の違いは遺伝子のどのような違いによって生み出されるのでしょうか。現在研究を進めているところです。
エキシンの穴の中にはポレンコートと呼ばれる粘着性の物質が存在します。ポレンコートには雌しべが同種植物の花粉かどうかを識別するための目印が含まれているようですが、その実体はまだよくわかっていません。それはどのような物質なのか、また、ポレンコート自体がどのようなしくみで作られるのかについても研究しています。
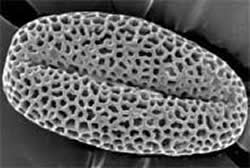

(図2)シロイヌナズナの花粉の走査型電子顕微鏡像。野生型(左)の花粉は網目状構造のエキシンで覆われているが、kaonashi9 突然変異体の花粉のエキシンは突起状である。1個の遺伝子の機能が失われるだけでエキシンの構造は大きく変化する。
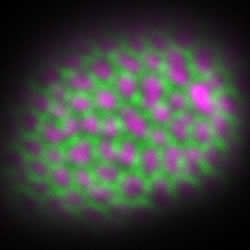
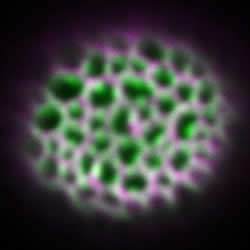
(図3)エキシンが形成され始めた時期のシロイヌナズナの若い花粉の表面に存在するペクチン(左図の紫)とアラビノガラクタン(右図の紫)を免疫染色法で可視化した。緑はオーラミンで染色されたエキシン。ペクチンが網の目の中に存在するのに対し、アラビノガラクタンは網目状のエキシンの外側に貼り付いている。実際にはペクチンやアラビノガラクタンの構造が先に作られ、その隙間にスポロポレニンが集積してエキシンが成形されるのだと考えられる。
遺伝子の糖応答性発現制御機構の解明
イモや種子に栄養が蓄積されるしくみ
植物は、光合成のエネルギーを使って作った栄養分を、次世代のためにイモや種子などの貯蔵器官に運んで蓄える性質があります。葉で作られた光合成産物は主にショ糖の形で篩管中を運ばれ、イモや種子に到着すると、そこでデンプン、油脂、タンパク質などに変換されて貯蔵されるのです。このとき、イモや種子では、運ばれてきたショ糖に応答して、デンプンや油脂を生合成する酵素の遺伝子や、貯蔵タンパク質の遺伝子の発現が活性化されます。この遺伝子発現の変化はどのようなしくみで起きるのでしょうか?糖によって活性化される転写制御因子を同定し、その性質を調べています。
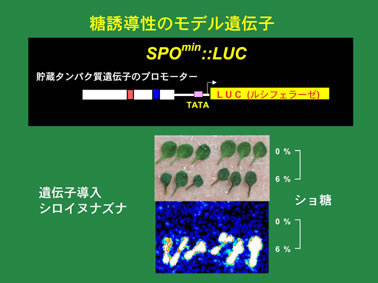
(図4)サツマイモの貯蔵タンパク質スポラミンの遺伝子のプロモータには、糖に応答して転写を活性化させるような転写制御領域が存在する。その領域(SPOmin)にルシフェラーゼ遺伝子(LUC)をつないでシロイヌナズナに導入した。下図は、この遺伝子を持つシロイヌナズナの葉を0%および 6%ショ糖で処理し、ルシフェラーゼの基質であるルシフェリンをかけて、発光の様子を高感度カメラで撮影した写真。6%ショ糖で処理した葉のみでルシフェラーゼが作られ、ルシフェリンが分解されて、発光している様子がわかる。