

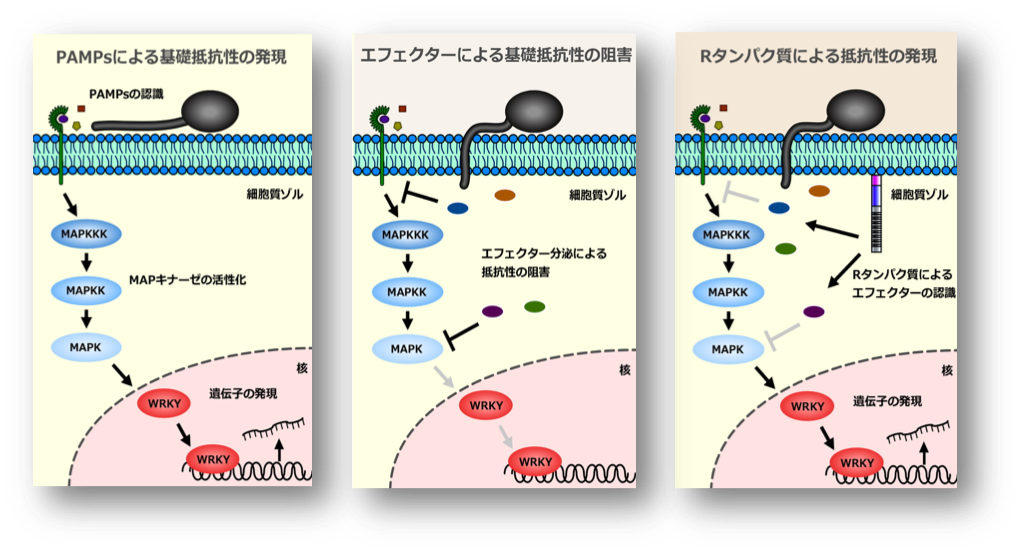
植物免疫の仕組み。
植物は、細胞表層や内部に病原菌を感知するアンテナである様々なレセプターを持っており、侵入者を認識し、防御反応を誘導します。このレセプターの構造が動物の自然免疫を担うレセプターと似ていることから、植物の生体防御機構は「植物自然免疫」と呼ばれようになりました。
植物は、菌類、細菌などの病原菌の攻撃に対して主に2つの認識機構を通じて病原菌を排除する植物独自の免疫機構を備えています。1つは比較的弱い抵抗性であり、鞭毛タンパク質フラジェリン、キチン、EF-Tuやリポ多糖など、病原菌の極ありふれた構造である病原体由来分子パターン (pathogen-associated molecular patterns; PAMPs) に対応する受容体が誘導する基礎抵抗性 (basal defense) です(図左)。病原菌は、自身の感染を成立させるために、エフェクター分子を植物細胞内に挿入し、基礎抵抗性を抑制します(図中央)。これに対して、植物は、病原菌の病原性因子であるエフェクタータンパク質をRタンパク質によって認識し、動的防御反応として過敏感反応 (hypersensitive response; HR) を伴う強固な抵抗性を示します(図右)。さらに病原菌は、エフェクターの構造を変異させて植物の抵抗性を回避しようとします。このように、植物と病原菌は、短いスパンでお互いを進化させてきました。しかし、いずれの抵抗反応も、MAPキナーゼカスケードにより制御されると考えられています。
植物免疫学は、植物と病原菌の間で展開される分子の相互作用を明らかにする分野として捉えることができます。
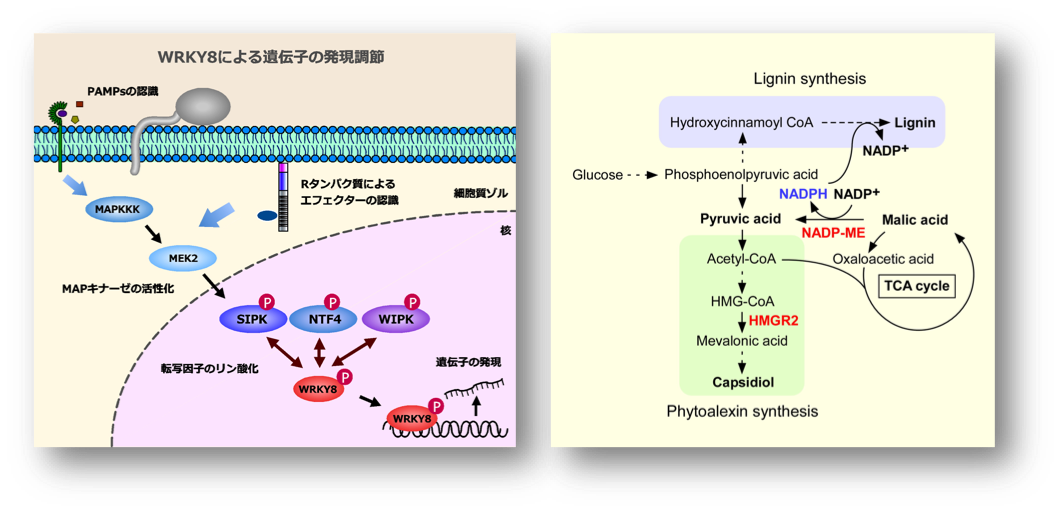
MAPキナーゼによる免疫調節機構。
MAPキナーゼカスケードは、真核生物に高度に保存されたシグナル伝達経路で、MAPKKK (MAPキナーゼ・キナーゼ・キナーゼ)、MAPKK (MAPキナーゼ・キナーゼ)、MAPキナーゼという3つのタンパク質リン酸化酵素で構成されています。近年、MAPキナーゼカスケードが、植物免疫応答の中枢を担うことが明らかになってきました。タバコでは、病害シグナルに応答してWIPK (wound-induced protein kinase)、SIPK (salicylic acid-induced protein kinase) およびNTF4が、MAPKKであるMEK2によりリン酸化され活性化します。MEK2の恒常的活性型変異体であるMEK2DDを一過的に発現させると、活性酸素・活性窒素の生成にはじまり、防御関連遺伝子の発現誘導、HR様の細胞死が誘導されます。
私達の研究室では、MAPキナーゼカスケードによる免疫誘導機構を明らかにする目的で、SIPKの基質タンパク質を明らかにしてきました。これまでに、ジャガイモのSIPKによりリン酸化されるタンパク質として、9個の基質を同定しました。これらの中で、植物特有の転写因子であるWRKY8は、SIPK、WIPK、NTF4により細胞核内でリン酸化されることを明らかにしました。
リン酸化されたWRKY8は、DNAとの結合活性を高め、標的遺伝子の発現を誘導することが解りました。WRKY8は、リン酸化されることにより、リグニン合成に関与すると考えられているNADP-malic enzyme (NADP-ME) 遺伝子や、ファイトアレキシン合成の鍵酵素である3-hydroxy-3-methylglutaryl CoA reductase (HMGR) 遺伝子を正に制御することを突き止めました。すなわち、病原菌を感知した植物は、リン酸化反応系を通じてSIPK、WIPK、NTF4という3つのMAPキナーゼを活性化します。これらによってリン酸化されたWRKY8は、抗菌物質であるファイトアレキシンの合成酵素遺伝子の発現を誘導してファイトアレキシンであるカプシジオールを生成させ、免疫応答を亢進することを発見しました。
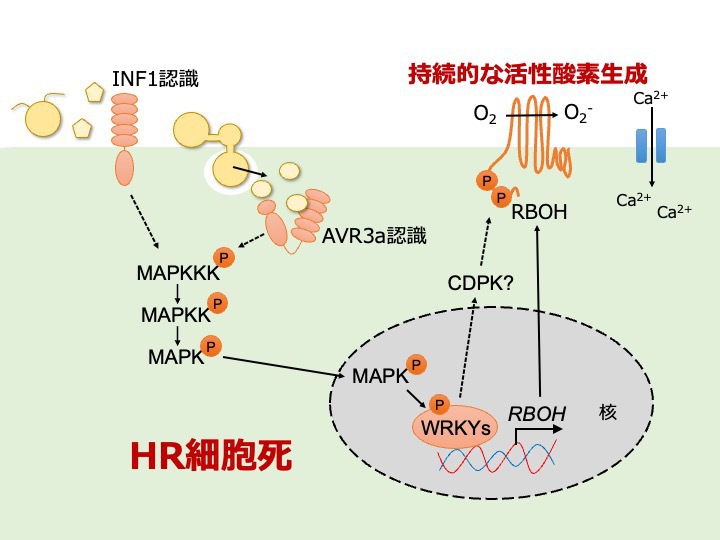
ETIにおける活性酸素生成NADPHオキシダーゼの制御機構。
ETIにおける持続的な ROS バーストには,MAPK-WRKY 経路を介した NbRBOHB の転写導が必要であることを明らかにしました。