TOP >> �������e >> �����O���[�v
���݁A�ȉ��̂悤�ȃe�[�}�Ō�����i�߂Ă��܂��B
�����O���[�v
(1) �C�l�ȐA���̍��̒ʋC�g�D�`���@�\�̉�
�g�E�����R�V�Ȃǂ̃C�l�ȐA���ł́A�y��ɉߏ�Ȑ������܂܂ꂽ��Ԃō͔|����ƍ��̔�w�Ɂi�j���j�ʋC�g�D���`������܂��i�}1-1�j�B
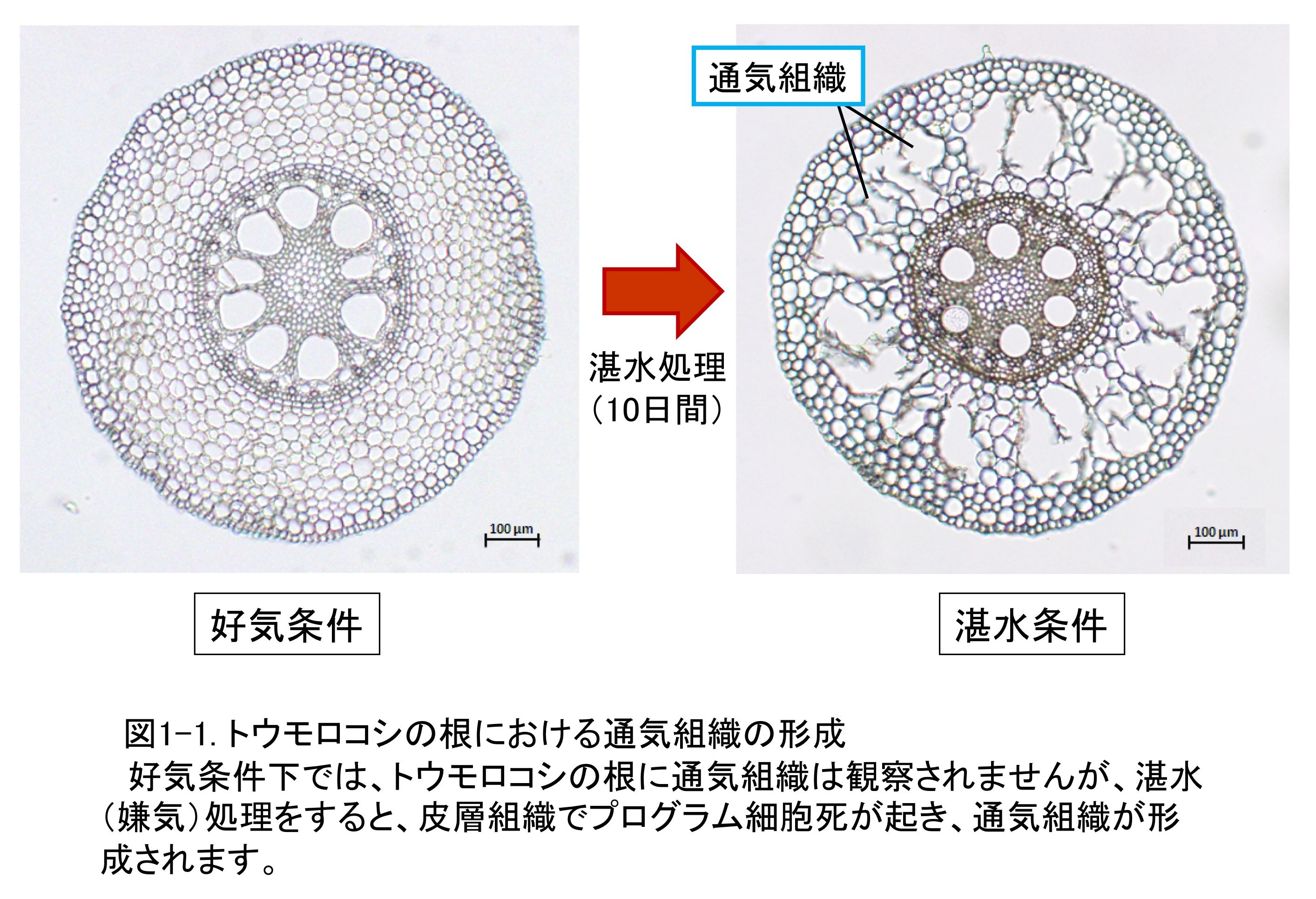
�ʋC�g�D�̎�Ȗ����́A���̊�����[���ւƎ_�f���������邱�Ƃł���A���̔��B���x���ώ����̋��������߂�v���̈�ƂȂ�܂��B�C�l�ȐA���̒ʋC�g�D�́A��w�g�D���ٓI�ɍזE�����邱�Ƃɂ���Č`������邱�Ƃ���A��`�q�ɂ���Đ��䂳�ꂽ�v���O�����זE���ł���ƍl�����Ă��܂��B�v���O�����זE���͈�`�q�̔��������ϋɓI�ȍזE���ŁA�A���ł͓��nj`���A�A�����[�����w�̏����A�q�t��̗�J�Ȃǂ̊튯�`���ɂ�����זE����A�a���ۊ����̍ۂɋN����ߕq���זE�����悭�m���Ă��܂��B�����̃v���O�����זE���ł́A���̈��ł���A�|�g�[�V�X�ɂ��Đ���Ɍ������s���A���̕��q���J�j�Y���̑�v�����炩�ɂȂ��Ă��Ă��܂��B���̈���ŐA���̃v���O�����זE���̕��q�@�\�ɂ��Ă͖����s���ȓ_�������Ƃ����̂�����ł��B �������́A�g�E�����R�V��C�l�̍��̒ʋC�g�D�`���ɂ�����v���O�����זE���ɒ��ڂ��A���ꂪ�A���̑ώ����Ɋ�^����@�\�Ɋւ���m���邱�Ƃ�ړI�Ƃ��Č������s���Ă��܂��B

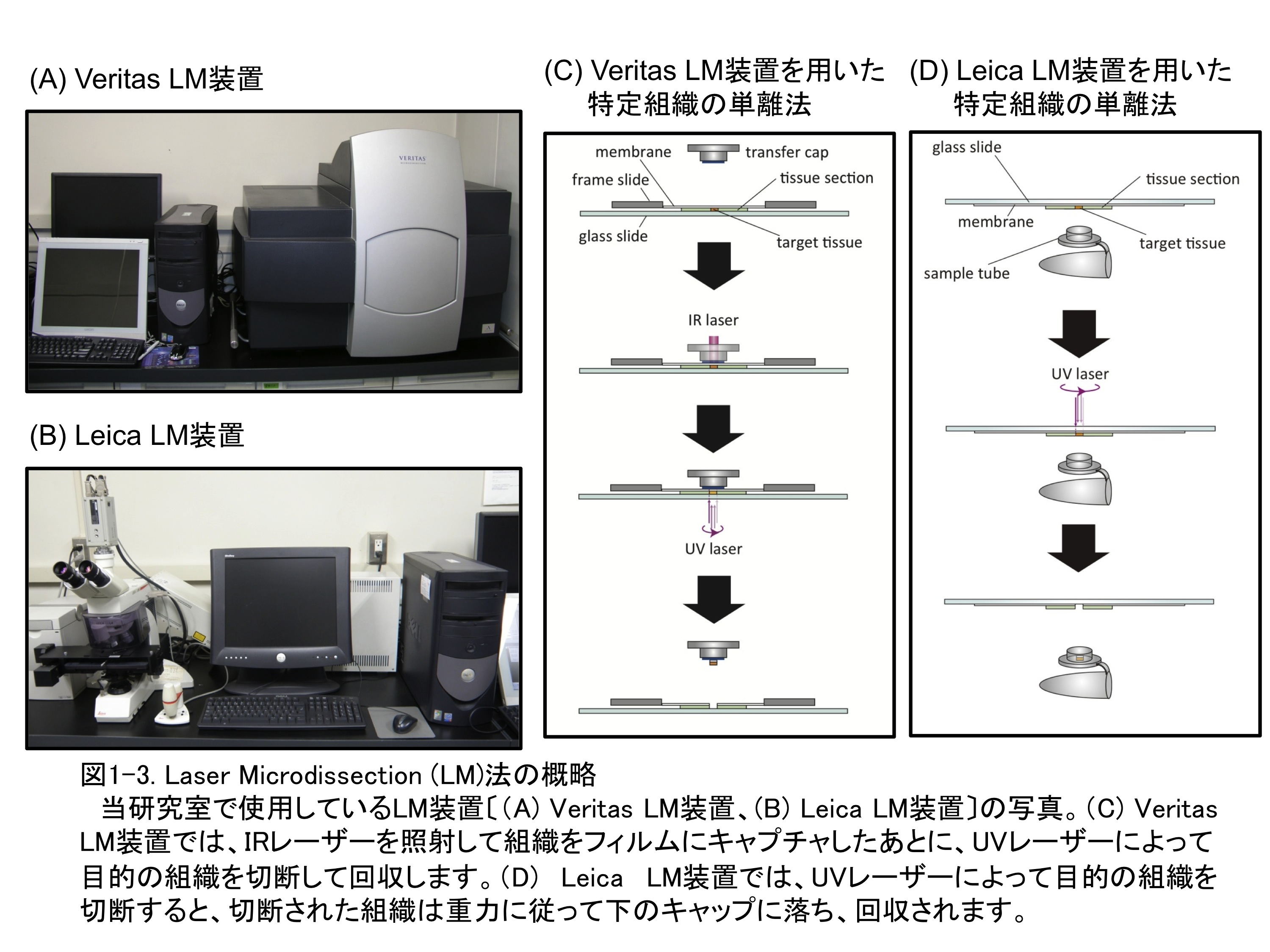
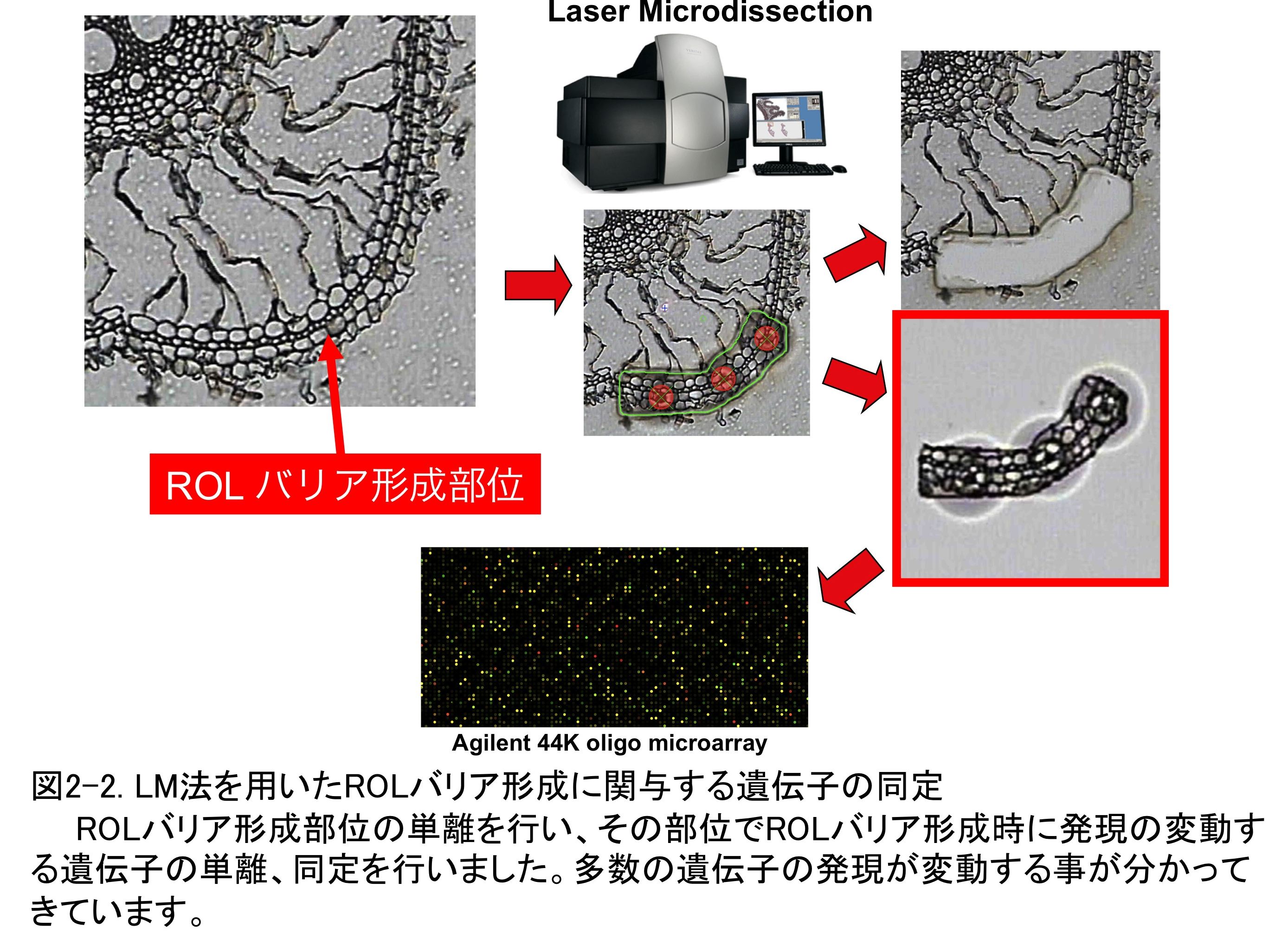
�g�E�����R�V�̍��̒ʋC�g�D�`���Ɋւ��v���O�����זE���́A��w�g�D�œ��ٓI�ɋN���܂��B���̈���ʼnߎ������ɂ���_�f�X�g���X�͔�w�g�D�����ł͂Ȃ����̑g�D�S�̂ɉe�����y�ڂ��A�L�͂Ȉ�`�q�̔����ʂ�ω������邱�Ƃ��\�z����܂��B���������č��̑g�D�S�̂�ΏۂɈ�`�q������͂��s�����ꍇ�A��w�g�D�ȊO�̑g�D�Ŕ��������`�q���܂܂�邱�Ƃɂ��A�T���̐��x���ቺ���Ă��܂��\�����l�����܂��B�����ŁALaser Microdissection�iLM�j�@�𗘗p���āA��w�g�D�݂̂�P�����邱�Ƃɂ��܂����i�}1-2�j�BLM�@�Ƃ́A�ؕЂɂ����W�{�̓��蕔�ʂ̎��͂Ƀ��[�U�[�������ĂĐ؏����A�ړI�̑g�D�������ɒP�����邱�Ƃ��\�ȋZ�p�ŁA�P�������g�D���璊�o����RNA��^���p�N����p���đg�D���ٓI�Ȕ�����͂��s�����Ƃ��ł��܂��i�}1-3�j�BLM�@�ŒP��������w�g�D����RNA�𒊏o���āA�}�C�N���A���C��͂��s���A�ʋC�g�D�`���Ɋ֗^����ƍl�������`�q�𑽐����肵�܂����B
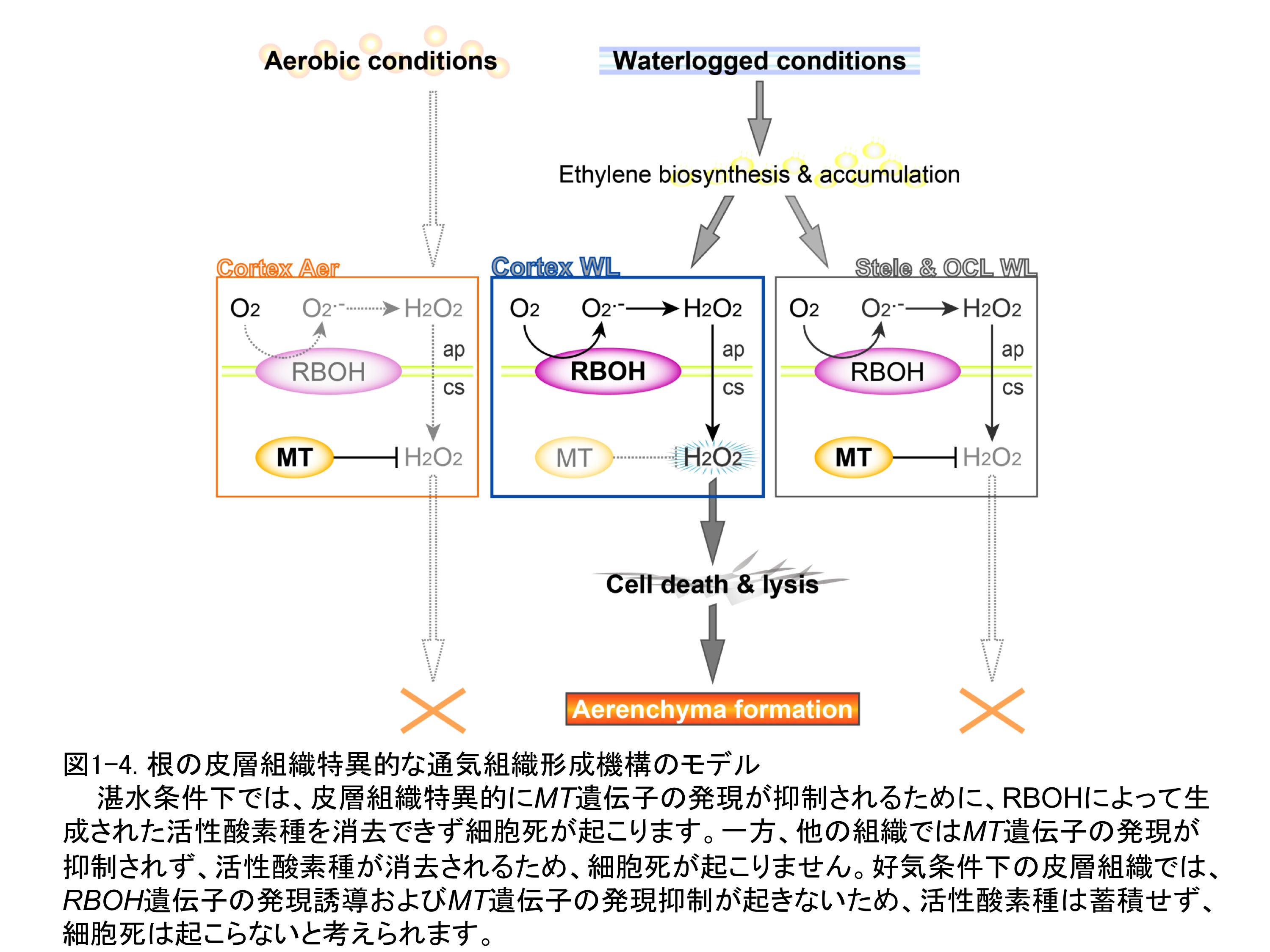
���̌��ʁA�ʋC�g�D�`���ߒ��ŁA�����_�f��iO2.-�j�����̌��y�fNADPH oxidase�̐A���z�����O�ł���Respiratory burst oxidase homolog�iRBOH�j��`�q�̔������U������A�a�Q�����ɂ����Ċ����_�f��iH2O2�j�̏�����S�����Ƃ�����Ă���Metallothionein�iMT�j��`�q�̔������}������邱�Ƃ����炩�ɂȂ�܂����B����ɁALM�@�ŒP���������S���A��w�A�\��E�O����܂ފO�w�ɂ����Ĉ�`�q������͂��s�������ʁARBOH��`�q�̔����U���͖��m�ȑg�D���ِ��������Ȃ������̂ɑ��āAMT��`�q�̔����͔�w�g�D���ٓI�ɗ}������邱�Ƃ�������܂����B�����̌��ʂ��A��w�g�D�ł�MT��`�q�̔������}�����ꂽ���ʁARBOH�ɂ���Đ������ꂽ�����_�f��������ł��Ȃ��Ȃ�זE�����N����̂ɑ��āA���̑g�D�i���S���A�\��E�O����܂ފO�w�j�ł�MT��`�q�̔����}���������Ȃ����߂ɁAMT�ɂ���Ċ����_�f�킪���₩�ɏ�������A�זE�����N���Ȃ��Ƃ������f������܂����i�}1-4�j�B���݁A���̃��f���̌���i�߂Ă��܂��B
(2) �C�l�̍�����̎_�f�R�o��h���o���A�iROL�o���A�j�̌`���@�\�̉�
���앨���ߎ����ŏ�Q�������錴���̈�ɁA���[�Ɏ_�f�������悭��������Ȃ����߂ɍ��̊��͂������邱�Ƃ�����܂��B��ʓI�ɁA�R���M�Ȃǂ̔��앨 �́A�C�l�Ɠ��l�ɁA�_�f��A�����邽�߂̒ʋC�g�D�����Ɍ`�����邱�Ƃ��ł��܂����A�_�f���ʋC�g�D����č��[�ֈړ�����ߒ��ŁA���̑啔�����ʋC�g�D���Ƃ�͂ޕ\��E�O�炩��y��֘R�o���Ă��܂��܂��i���̎_�f�R�o��Radial O2 Loss�iROL�j�ƌĂ�ł��܂��j�i�}2-1�j�B
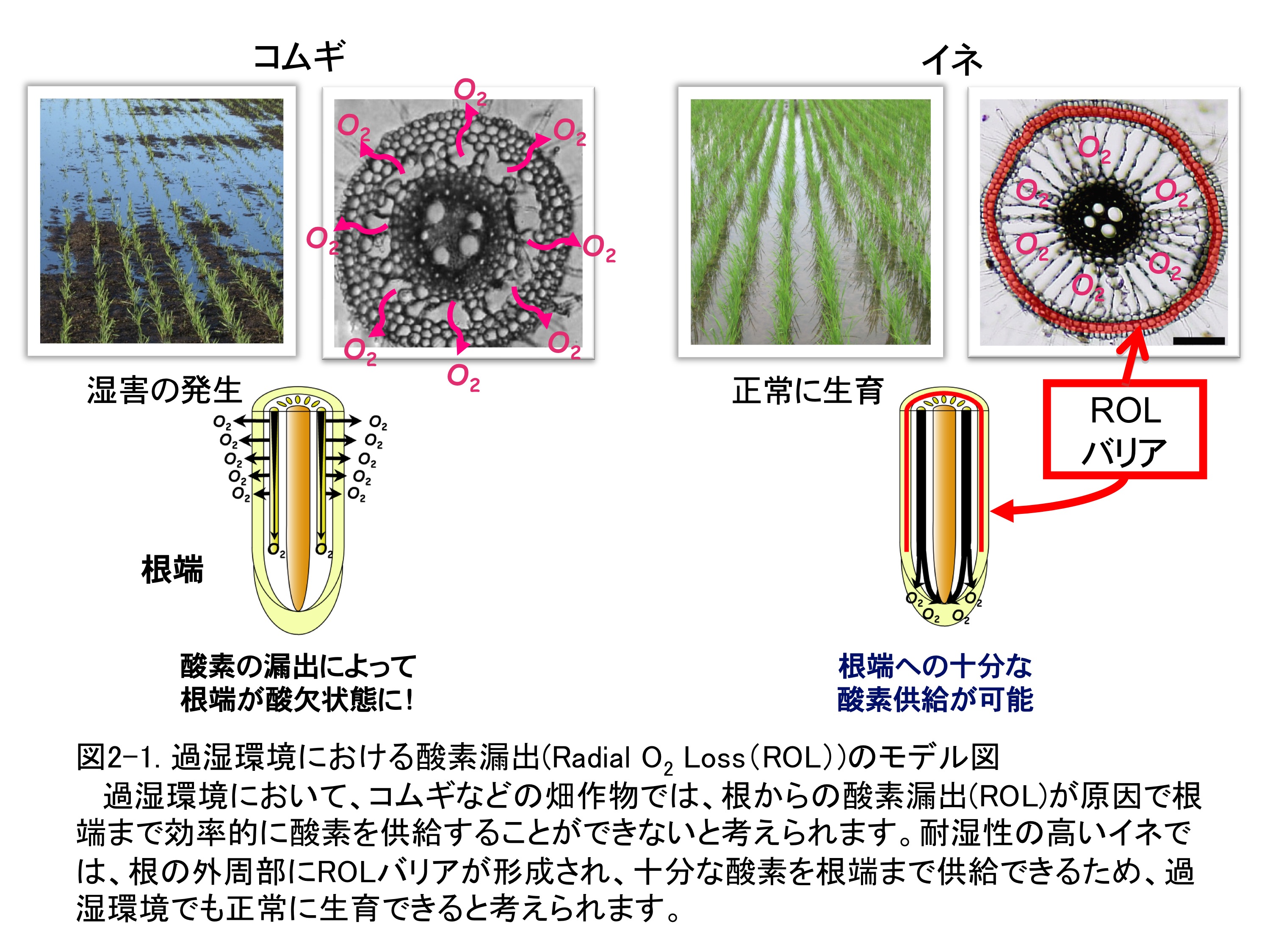
���̌��ʁA�����_�̂��鍪�[�����������_�f�ʂ��ɒ[�Ɍ������邽�߁A���̐L������~���A��Q�������Ă��܂��܂��B����A�ώ����̍����C�l�́A���̊�̒ʋC�g�D�̊O���ɁA�ߎ����ɗU���I�Ɏ_�f�̘R�o��}������o���A�iROL�o���A�j���`�����邱�Ƃɂ���āA���[�܂Ō����悭�_�f���������A�ߎ��y��ō���L�������邱�Ƃ��ł��܂��i�}2-1�j�B���̂悤�ɐA���̑ώ����ɂ͍��[�ւ̎_�f�^�����K�{�ł���܂����A����ɂ͒ʋC�g�D�̌`�������ł͏\���łȂ��AROL�o���A���`�����č��[�܂Ō����I�Ɏ_�f���������邱�Ƃ��d�v�ł���Ƃ����܂��B
���̂悤�ɁAROL�o���A�͑ώ����ɂ����ďd�v�Ȍ`���ł���ɂ��ւ�炸�A���o���@�̔ώG���Ȃǂ���A���̔��������40�N���o�߂������݂ł����� �\��������`���Ɋւ���`�q�͓��肳��Ă��炸�A���̂قƂ�ǂ����𖾂̂܂܂ł��B�������́A����ROL�o���A�`�����ʂ�LM�@�ɂ���ĒP�����āA�}�C�N���A���C��͂��s�����Ƃɂ��ROL�o���A�`�����ɊO����ӑg�D���ٓI�ɔ��������`�q�𑽐����肵�܂����i�}2-2�j�B���݁A������`�q�̋@�\��͂��s���AROL�o���A�`���Ɋւ���`�q�̓�������݂Ă��܂��B
�����O���[�v
(3) �_�C�Y�̓ʋC�g�D�̌`���@�\�̉�
���{�̃_�C�Y�͔|�ɂ����āA8���ȏオ���c�]�����ōs���Ă��܂��B���c�́A���߂�K�v�����邽�߁A��{�I�ɔr�����������y��ł��B���̂��߁A���앨�ł���_�C�Y�𐅓c�]�����ō͔|����Ǝ��Q�������A���ʂ̒ቺ�������Ζ��ɂȂ�܂��B���̂��߁A�_�C�Y�̑ώ�������͏d�v�Ȕ_�Ɖۑ�ƂȂ��Ă��܂��B
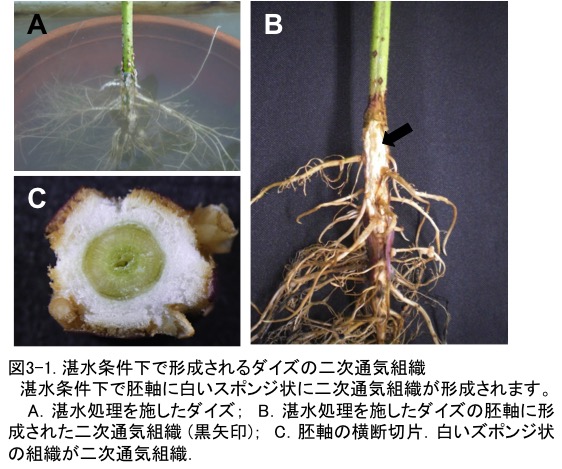
�_�C�Y�̒X���y��ւ̓K���`���̈�ɓʋC�g�D�̌`��������܂��B�X���������{�����_�C�Y�́A�n�㕔����n�����֎_�f���������邽�߂ɁA�A���A�����Ƃ������튯�ɐV���ɒʋC�g�D���`�����܂��i�}3-1�j
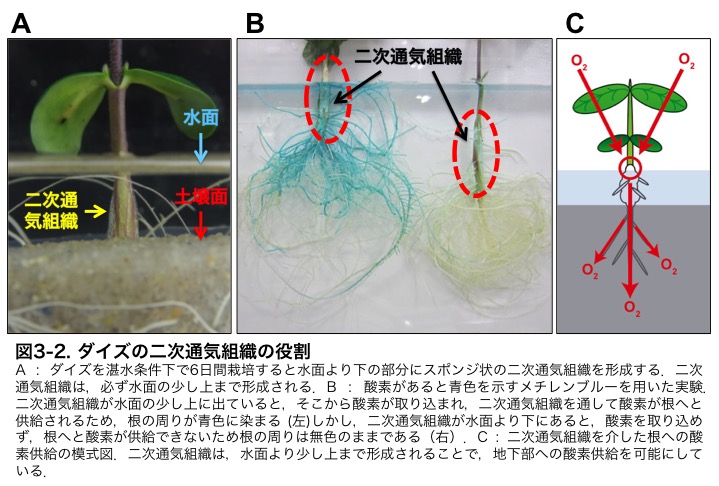
����́A�C�l��g�E�����R�V�����̔�w�g�D�Ƀv���O�����זE���ɂ���Č`������ꎟ�ʋC�g�D�Ƃ͈قȂ�A��w�g�D�̓����ɐV���Ɍ`������邱�Ƃ���ʋC�g�D�ƌĂ�Ă��܂��B���̓ʋC�g�D�́A���ʂ̏�����܂Ō`������邽�߁A�_�f�����̓�����Ƃ��Ĕ��ɏd�v�ȑg�D�ɂȂ�܂��i�}3-2�j�B
���̂悤�ɁA�ʋC�g�D�������w�I�ɂǂ̂悤�Ȗ����������Ă��邩�A�܂��`�Ԋw�I�ɂǂ̂悤�ɔ������邩�͕���Ă��܂����B�������A�ʋC�g�D���ǂ̂悤�ȃ��J�j�Y���ɂ�蔭�����邩�͑S���m��������܂���ł����B�����ŁA�������͂��̓ʋC�g�D�̌`�����J�j�Y�����𖾂��邽�߂ɉ�͂��s���Ă���܂��B
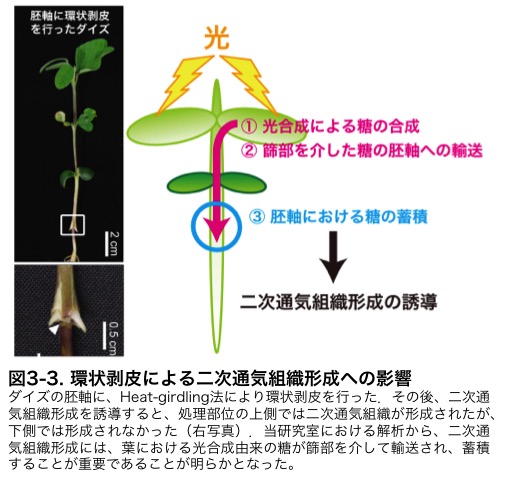
�������́A����܂łɈÏ��ł͓ʋC�g�D���`������Ȃ����Ƃ����A��������A�������Y���ł���V�������d�v�ł��邱�Ƃ𖾂炩�ɂ��܂����B�����[�����ƂɁA��Heat-girdling�ƌĂ���@�ɂ�����{���ؕ��g�D�ȊO��������ƁA�����������ʂ̏㑤�ł̂ݓʋC�g�D�̌`�����U������邱�Ƃ��킩��܂����i�}3-3�j�B����ɉ����l�X�Ȏ������ʂ���A�t�ɂ�����������R���̃V�������A⿕�����ėA������邱�Ƃ��ʋC�g�D�̌`���̂��߂ɏd�v�ł��邱�Ƃ����炩�ƂȂ�܂����D���݁A����Ȃ�ڍׂȃ��J�j�Y���𖾂炩�ɂ��邽�߂ɁA��`�q������͂Ȃǂ𒆐S�ɍs���Ă��܂��B
(4) �g�}�g�ɂ�����ؕ��g�D�̔��B�@�\�̉�
�A�������̗ǂ����������������ɂ́A�������Y���̓]�������ł͂Ȃ��A�{�����̋������K�v�s���ł��B�������̌������ł́A���ʐ��̈قȂ�g�}�g�i��A�ƕi��B���r�����Ƃ���A�W���i��A�ɔ�ׁA�����i��B�ł́A��s�ɂ�����ؕ��g�D��10%���x�傫�����B���Ă��邱�Ƃ��킩��܂����i�}4-1�j�B�ؕ��g�D�́A������z�������{������t��ʎ��Ȃǂɋ�������p�C�v�Ƃ��Ă̖������ʂ����Ă��܂��B�����ŁA�������͂��̖ؕ��g�D�̔��B�̍�����̕i��̎��ʐ��̍��ɉe�����y�ڂ��Ă���̂ł͂Ȃ����ƍl���A�������s���Ă��܂��B���̌�����ʂ��āA���i���E�����̃g�}�g�i��̍�o��ڎw���Ă���܂��B
�@
(5) �C�l�̓S�ߏ�ϐ��@�\�̉�
�@�{�����́A�C���h�l�V�A�̍^����P�n�ɂ�����앨���Y�̈��艻�Ɍ����A��肪���������Ă��銥���A������ю_���y��ɂ���Q���y�����邱�Ƃ�ڎw���������v���W�F�N�g��1�e�[�}�Ƃ��Đi�߂Ă��܂��B�����I�ɁA�R�����Y�̐���ȃC���h�l�V�A�����ɍL���鐶�Y�|�e���V��������������n��ΏۂɁA�O�q�̃X�g���X�ϐ��i��̑I�肨��уX�g���X�ϐ��@�\�̉𖾂�ʂ��āA�R���̐��Y���艻�ɍv���������ƍl���Ă��܂��B
�C���h�l�V�A�̑����̏���n�ł́A�_���y��������ł̓S�ߏ�X�g���X������͔|�̐[���Ȗ��ƂȂ��Ă��܂��B�����Ŏ������́A�S�ߏ�ϐ��C�l�i��̑I�����A�ϐ��ɂ������L�pQTL�肵�A�C���h�l�V�A�̎�͕i��ł���Ciherang�ɓ������邱�ƂœS�ߏ�ϐ�������Ciherang����o���A�C���h�l�V�A�̃R�����Y�ɍv���������ƍl���Ă���܂��B
�A���ɂ�����S�ߏ�̏Ǐ�Ƃ��āA�n�㕔��n�����̐���j�Q�͂������A�t�ȂǂɃu�����W���O�ƌĂ��Ԋ��F�̕a��������邱�Ƃ��悭�m���Ă���܂��i�}�T�|�P�j�D

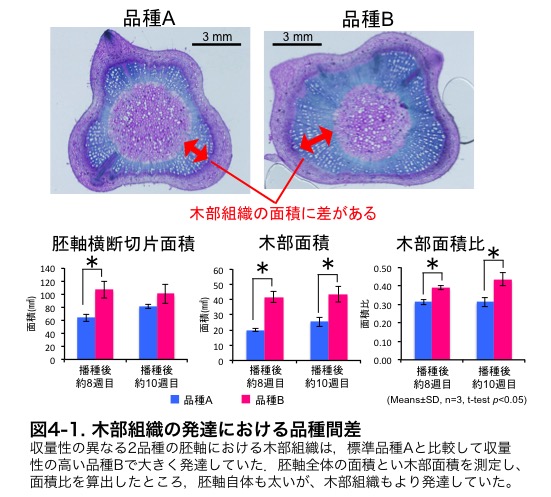
�����Ŏ������̌������ł́A�������̃C�l�i��̒�����S�ߏ���ł�����j�Q��u�����W���O���ώ@����Ȃ��i��A�ƕi��B��I�����܂����B�i�}�T�|�Q�j�D

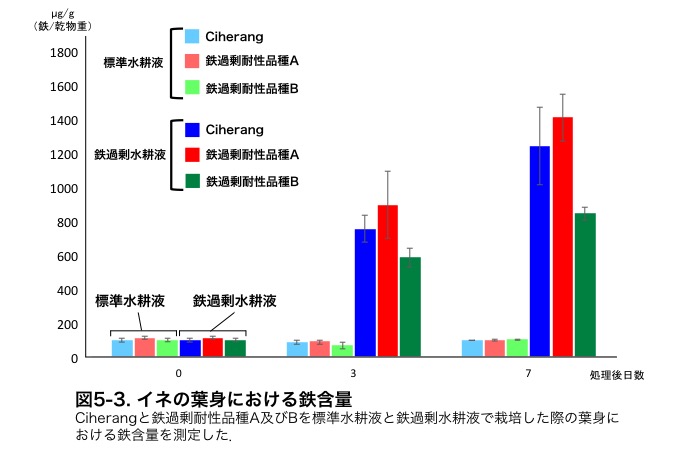
�����̕i��̃C�l�̗t�ɂ�����S�ܗʂ𑪒肵���Ƃ���A�S�ߏ萅�k�t�ō͔|�����ۂɁA��ϐ��i��ł���Ciherang�ƓS�ߏ�ϐ��i��A�̊Ԃł͑傫�ȍ��͂���܂���ł������A�S�ߏ�ϐ��i��B�ł͗L�ӂɓS�ܗʂ��ቺ���Ă��܂����i�}5-3�j�D�����̂��Ƃ���S�ߏ�ϐ��i��A��B�őϐ��@�\�ɈႢ������\�����l�����܂����D
���݂����Q�̕i���p����QTL��͂��s���S�ߏ�ϐ��Ɋւ��QTL�̓�����s���Ă��܂��D
�p�O���[�v
�������e�̊ȒP�ȏЉ���������̋L���ŏЉ�Ă��܂��D
�܂��C�������e�̓����Youtube�ŏЉ�Ă��܂��D
(6) �͔|�C�l��p������^����Ɋւ���`�q�̒T���Ɨ��p
�@�C�l�̕�͑����̎}������(���})����\������镡�G�ȍ\�������Ă���A�C�l�̎��ʍ\���v�f�̈�ł��钅�����͕�̕��}�p�^�[���̐���ɂ���Ă����炳���ƍl�����܂��B�������́A�����C�l�i��Ɍ������̕��}�\���ɒ��ڂ��A��^�̑傫���قȂ�i��Ԃ̌��G����p����QTL��͂ɂ�蕪�}�p�^�[���𐧌䂵�Ă���L�p��`�q�̒T����i�߂Ă��܂��B
(7) �쐶�C�l��`�����̑��l���𗘗p�����A���ɐB�헪�̗���
�@�쐶�C�l�͑��l�Ȋ��ɐ������ɐB���œK�����Ă������߁A���̌`�ԓI�E��`�I���l���͐A���̔ɐB�헪�𗝉�����̂ɓK�����ޗ��ł��B���Ɏq���̐��ɑ傫�ȉe����^�����`�Ԃ́A�ɐB�̍œK���ɍł��d�v�ȗv�f�ł���A���̑��l�����x�����`��Ղ��𖾂��邱�ƂŁA�쐶��̔ɐB�헪�̑��l���𗝉��ł���ƍl�����܂��B�����Ŏ������́A�쐶��`������������`�Ԃ̑��l���𗘗p���āA��`�ԑ��l����`�q�̒P���Ƃ��̋@�\��͂�i�߂Ă��܂��i�}7�j�B

![]()