
| トップ | 研究概要 | メンバー | 研究発表 | 就職状況 | 写真集 |
C4植物の葉緑体の細胞内配置に関する研究
イネ科植物の葉構造
私達がふだん何気なく見かける植物もその構造や機能は複雑で多様です。
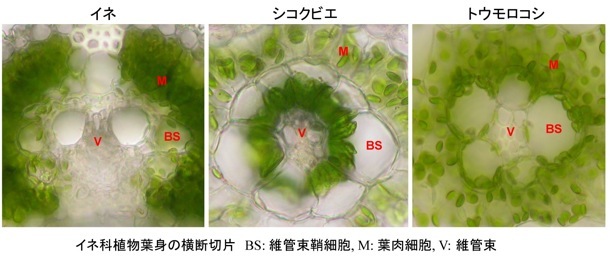
下の写真は、主要作物であるイネと雑穀のシコクビエおよびトウモロコシの葉身横断切片の光学顕微鏡像です。どの植物もイネ科に属し、葉脈が平行に並んだ細長い葉をもち、細胞の基本的な配置は似ています。水や栄養分の通り道である維管束(V)の周りを一層の維管束鞘細胞(BS)が取り囲み,さらに外側を葉肉細胞(M)が取りまいています。
各細胞内の緑色の顆粒が葉緑体です。
イネでは葉肉細胞にぎっしりと葉緑体が詰まっている一方、維管束鞘細胞にはごく僅かの葉緑体しか存在しません。イネは葉肉細胞の葉緑体で主に光合成炭酸固定(カルビン・ベンソン回路)を行っており、C3植物に分類されます。
シコクビエの緑葉横断面はイネとは少々様子が異なっています。イネに比べて葉肉細胞に存在する葉緑体の数は少なく、細胞内に散在しています。その内側に位置する維管束鞘細胞の葉緑体はイネに比べて大型で数が多く、中心の方(維管束側)に片寄っています。シコクビエではこの両細胞にまたがってC4光合成回路が機能しており、C4植物とよばれます。
トウモロコシもC4植物ですが、維管束鞘細胞の葉緑体はシコクビエとは異なり、維管束側から遠い方、すなわち葉肉細胞側に偏って配置されています。
環境ストレスに応答した葉肉葉緑体の凝集運動
隣り合う葉肉細胞と維管束鞘細胞が明白に分化しているC4植物では、両細胞で機能している酵素タンパク質が顕著に異なり、葉緑体の細胞内配置も異なっています。私達はこの葉緑体の細胞内配向性に興味をもち、配向性に関わるメカニズムや生理的意義を解明しようとしています。
今まで両細胞の葉緑体配置は常に一定だと考えられていましたが、シコクビエの葉に極強光を照射すると、細胞膜に沿って偏在していた葉肉葉緑体が維管束側に集合することを私達は見つけました。過剰な光を受けると葉緑体中に活性酸素が生じ、光合成活性の低下(光阻害)、葉緑体タンパク質の失活・分解,膜構造の崩壊が引き起こされてしまいます。葉緑体が集まることでより内側の葉緑体が受け取る光エネルギーが軽減されると考えると、葉肉葉緑体の集合運動は光阻害に対する防御機構だと推定できます。
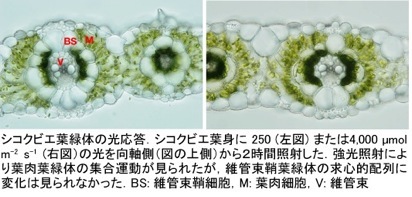
さらに、乾燥、塩、あるいは浸透圧ストレスを受けたシコクビエでは通常光下でも葉肉葉緑体の凝集運動が見られました。また、葉肉葉緑体の凝集運動は真夏の炎天下で生育するシコクビエでも観察されました。 過酷な環境に曝されている自然条件下で起こる現象だと考えられます。同様の現象はトウモロコシでも確認しており、C4植物に共通の現象である可能性があります。私達は、この凝集運動の生理的意味を解明することを目指しています。
維管束鞘葉緑体の細胞内配置の維持
一方、さまざまな環境ストレスを与えても維管束鞘葉緑体の配列性はビクともしません。ただ、葉片を遠心して葉緑体に遠心力をかけるとさすがにその配列性は崩れました。しかし、1〜2時間後には元の配列に戻ってしまいます。したがって、葉緑体の配向性を頑固に維持する機構が維管束鞘細胞特異的に存在し、葉緑体(あるいは葉緑体を動かす装置)は細胞内で自身の位置を認識していると考えられます。また、維管束鞘葉緑体が細胞内の一カ所に局在することは葉細胞が効率的に代謝活動を行う上で重要なのだとも推察されます。
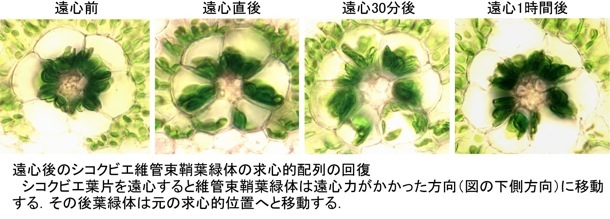
これらの両葉緑体の移動には細胞骨格のアクチン繊維が関与していることを阻害剤を用いた実験で確認していますが、配列の違いを引き起こすしくみを今後解明していく必要があります。一般にC4植物はC3植物に比べて環境耐性能が高いといわれますが、その要因として葉緑体の細胞内配列性が関係しているかもしれません。
本研究で得られる知見をもとにして、環境耐性能を向上させた作物開発のための新しいターゲットが提供できることを期待しています。また、葉緑体が細胞内で自身の位置をどのように認識し、方向性を持って移動しているのか、そのメカニズムも解明したいと考えています。