窒素代謝に関連する微生物群集
1)有機態窒素の無機化
2)硝化と脱窒
3)窒素固定
水田には洪水防止,水涵養,水質浄化,気温緩和,景観維持など多面的な機能30)が知られているが,最も重要な機能と役割はいうまでもなく,水稲の生産である。水田土壌の物理的・化学的・生物的な種々の作用や働きが水稲の生産を支えている。それらの水田土壌の働きの中で,無機養分としての窒素の供給や稲わら・堆肥といった有機物の分解など,土壌微生物の活動が重要な役割を果たしている場面は少なくない。
水田は,水稲の栽培期間中,湛水されることが,他の農耕地とは著しく異なる点である(図1)。湛水にともなって生じる作土の還元化と表面水(田面水)の存在は,他の耕地土壌には見られない水田土壌生態系の特徴であり,水田土壌で生じる物理的・化学的・生物的な作用だけでなく,生息する微生物群集にも大きな影響を及ぼす25)。
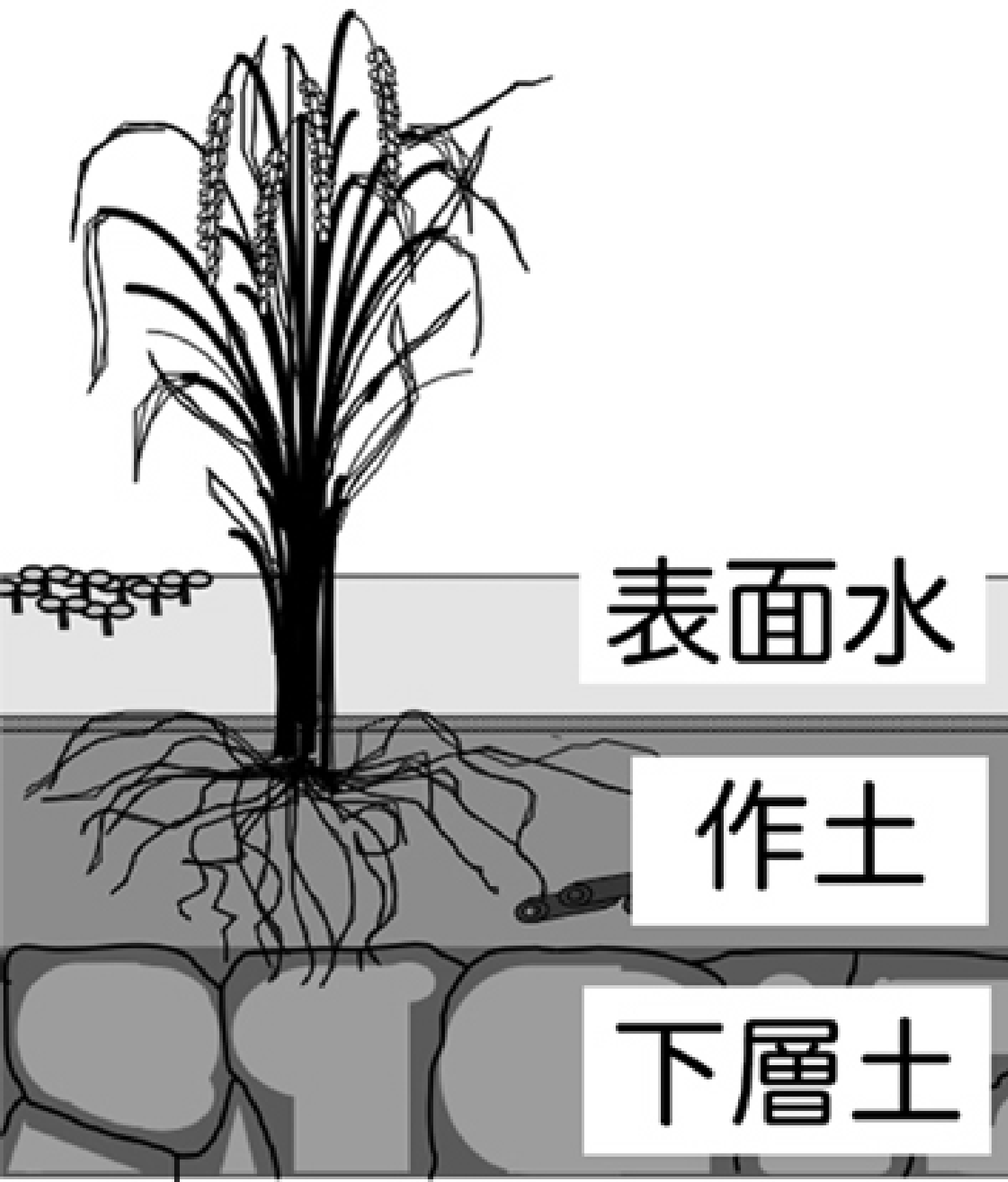
図1 水田生態系の構造(模式図)
ここでは,水稲生産を支える土壌の機能として,最も重要な無機養分である窒素の形態変化に注目し,水田土壌における窒素代謝に関連する様々な微生物群集の研究例を紹介する。それとともに,還元的な土壌中における特徴的な物質代謝として,有機物分解の最終過程に位置づけられるメタン生成を取り上げ,反応に関わる微生物群集について,これまでの研究事例を紹介する。これらの研究の中で,それぞれの代謝の鍵となる微生物群や,反応に重要な役割を果たしている微生物間の相互作用について,明らかにされた知見と未解明のまま残されている点をも含めて述べたい。
1)有機態窒素の無機化
土壌中の有機態窒素から生じた無機態窒素は,水稲が吸収する窒素量の半分程度を占めており27,44,50),地力窒素と呼ばれる。そのため,地力窒素の発現量と無機化パターンの把握は水稲の栽培・施肥管理にとって重要であり,古くから現在まで数多くの研究が行われてきた34)。地力窒素の主要な給源は,微生物バイオマスに由来するタンパク態と考えられている。したがって,土壌中のバイオマス窒素の無機化は,主として,タンパク質のプロテアーゼ分解によるアミノ酸生成とそれに続く,アミダーゼ,リアーゼ,オキシダーゼなどによるアミノ酸からのアンモニア生成とみなすことができる。このうち,第一段階を触媒するプロテアーゼが,硝化・脱窒を含むそれ以降の反応速度を律速する鍵酵素と考えられている14,15)。
このように,有機態窒素の無機化あるいは地力窒素の発現,特に第一段階のタンパク態窒素のプロテアーゼ分解は水稲生産にとって極めて重要な過程であるが,これらの代謝を担う微生物群集は全くといっていいほど研究されていない。唯一の例が,渡邊らにより行われた,土壌プロテアーゼ生産微生物に関する一連の研究56〜63)である。水田土壌のプロテアーゼの主な供給源が細菌であるとの知見13,16)に基づき,細胞外プロテアーゼ生産能の高い菌株の分離に有効なアゾコール平板培地59)を用いて,水稲栽培期間に水田土壌からプロテアーゼ生産性の細菌・放線菌の分離を行ったところ,得られたプロテアーゼ生産菌の大部分がBacillus属細菌であった56)。さらに,これらの分離株の中で,高い酵素生産能を持つBacillus属菌株のプロテアーゼの酵素学的特性が,水田土壌から抽出したプロテアーゼの酵素特性とよく一致したこと57,58),水稲栽培期間中の水田土壌におけるプロテアーゼ生産性のBacillus属細菌の動態とプロテアーゼ活性の変動の間に有意な相関が認められたこと60)より,水田土壌のプロテアーゼ生産を担う主要な微生物群がBacillus属細菌であると結論した。また,Bacillus属細菌の増殖時(栄養細胞)と胞子形成期で生産されるプロテアーゼのタイプが異なり,Bacillus属細菌の形態変化が土壌プロテアーゼ活性の制御機構の一つであると推定された62)。上述したように,水稲の吸収窒素量の半分程度を供給するため,水稲生産のために最も重要な水田土壌の機能といっても過言ではない地力窒素の発現に関し,その鍵微生物群を明らかにした極めて重要な知見である。
2)硝化と脱窒
酸化層,還元層およびその境界を含む水田表層部における硝化・脱窒現象は施肥窒素の肥効に関わる重要な代謝過程であり,塩入松三郎・青峰重範42,43)により発見され,深層施肥あるいは全層施肥法の開発へとつながったことは有名である。
硝化の第一段階はアンモニアから亜硝酸を生成するアンモニア酸化作用であり,主として独立栄養性のアンモニア酸化細菌により行われる17,26)。アンモニア酸化の第一段階(アンモニアのヒドロキシルアミンへの酸化)を触媒するアンモニアモノオキシゲナーゼ遺伝子(amoA)およびBetaproteobacteriaに属するアンモニア酸化細菌(NitrosospiraとNitrosomonas)を対象とした16S
rRNA遺伝子のそれぞれに特異的なプライマーが開発され,様々な環境中における独立栄養性アンモニア酸化細菌の群集構造が解析されている26)。愛知県安城農業技術センターの水田圃場あるいは同圃場の水田土壌を用いたポット試験・ミクロコズム実験より得られた試料(作土21),稲わら20),田面水39),土壌表層33,39),水稲根18),ミジンコ35))について,これらのPCRプライマーを用いたDGGE解析を行ったところ,既知の菌株とは類縁性が低い一部の断片以外は全てNitrosospiraに近縁であった(図2)。水田生態系では,Nitrosospiraに近縁なアンモニア酸化細菌が主要であることを示しており,水田以外の土壌生態系におけるこれまでの多くの研究結果と一致した17,26)。
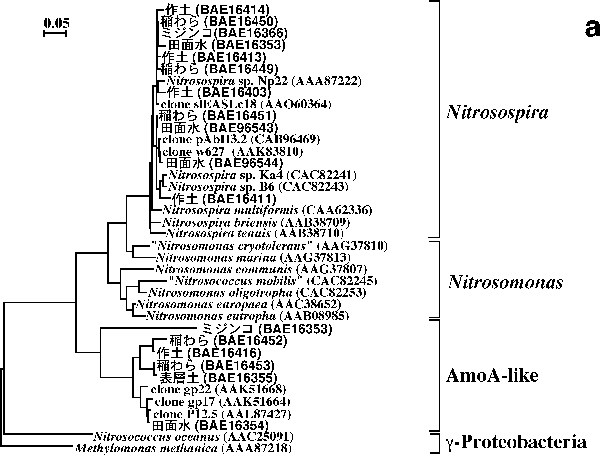
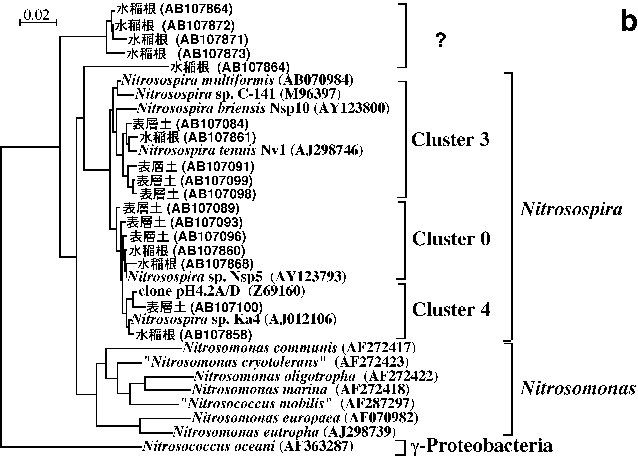
硝化の第二段階は亜硝酸の硝酸への酸化で,Nitrobacter, Nitrococcus, Nitrospina,
Nitrospiraが亜硝酸酸化細菌として知られている41,47)。これらはそれぞれ,alphaおよびgammaサブグループのProteobacetria門,Acidobacteriaに近縁な未確定の分類群(当初はDeltaproteobacteriaとされていた),Nitrospira門と,系統的に離れた分類群に属している41)。また,分離培養が難しく,分離例が少ないこともあり,環境中における亜硝酸酸化細菌の群集解析手法は一般的なものとなっていないが,最近になって排水処理系,水環境,底質などではNitrospiraの重要性が認識されつつある41)。培養法を用いた研究も含め,水田土壌に生息する亜硝酸酸化細菌群集はほとんど明らかにされていないが,水田の作土22)や植物遺体29)に生息する真正細菌全体を対象としたDGGE法による解析では,Nitrospiraに近縁な配列が得られている。水田土壌においてもNitrospiraが亜硝酸酸化細菌の主要な菌群であることが示唆されるが,さらにデータを蓄積する必要があろう。
脱窒は硝酸あるいは亜硝酸が酸素の代わりに呼吸のために電子受容体として利用され,一酸化窒素,一酸化二窒素(亜酸化窒素),分子状窒素へと還元される嫌気的な過程であり,細菌および糸状菌を含む広範囲の通性嫌気性微生物(脱窒菌)により行われる17)。各過程の酵素群とコードする遺伝子がPseudomonasなどの代表的な菌株を用いて明らかにされており17,65,66),これらの酵素遺伝子,特に亜硝酸還元酵素(Nir)や一酸化二窒素還元酵素(Nos)の特定のサブユニットをコードする遺伝子に対する特異的プライマーを用いて,様々な環境中の脱窒菌群集の解析が行われている5,8,48)。水田土壌の脱窒菌群集を解析した研究はこれまでのところ見当たらないが,これらの手法を用いた分子生態学的研究が今後進むものと期待される。一方,分離・培養法の改良により,畑下層土では,系統的に既知の脱窒菌とは類縁性の低い菌株を含む低栄養性の脱窒菌群が主要であったとの報告12)があり,水田土壌でも脱窒菌の培養・分離を併せて行う必要があろう。
3)窒素固定
水田の田面水と作土の最表層部には光合成窒素固定微生物が生息している23,55)。これは,耕地土壌における窒素固定菌の分布や生態面からみた場合,水田の特徴といえよう。中でもシアノバクテリア(ラン藻)による窒素固定が古くから注目され,水田土壌の窒素肥沃性維持に重要な役割を果たしていると考えられている3,51)。シアノバクテリアによる窒素固定量は一作当たり数〜80
kgN ha-1とされ38),九州農試(筑後市)の水田圃場における測定例では,約25 kgN ha-1の値が報告されている36)。培養や観察によって水田にはTolypothrix,
Anabaena, Nostoc, Calothrixなどのシアノバクテリアが生息することが明らかにされている53,54)が,このような生態学的研究よりはむしろ,接種や窒素固定量の把握と水稲への効果などシアノバクテリアの利用に重点を置いた研究が数多く行われてきた3,24,38,51,55)。最近,シアノバクテリアの16S
rRNA遺伝子に特異的なプライマーを用いたDGGE法により,水田作土のシアノバクテリアの群集構造が解析され,Subsection I〜IVに属するシアノバクテリアが得られている45)。水田の作土表層部に施用された稲わらを対象にして,同様の手法を用いて行った解析では,Subsection
IIIおよびIVの糸状体シアノバクテリアが主として検出された4)。
メタン生成は還元条件下の作土での有機物の嫌気的な最終分解過程として古くから研究されており,近年では温室効果ガスの発生過程としても注目されている2)。メタン生成は絶対嫌気性のメタン生成古細菌により行われる。水田土壌に生息するメタン生成古細菌としては,これまでに表1に示した属の菌株が分離されており,目(Order)としては,Methanobacteriales, Methanosarcinales, Methanomicrobialesに属する菌群であった1,10,31,32,49)。遺伝子を用いた解析では,これら3目に加えて,既知の培養菌の分類群と類縁性の低い,Rice cluster Iと呼ばれる配列群が得られており11,64),これらが水田土壌のメタン生成古細菌の主要な菌群であると考えられる。なお,ごく最近Rice cluster Iに属するメタン生成古細菌が培養・分離された19)。
表1 水田土壌から分離されたメタン生成古細菌の種類
| 目 |
属 |
| Methanobacteriales |
Methanobacterium |
| Methanobrevibacter |
|
| Methanosarcinales |
Methanosarcina |
| Methanosaeta |
|
| Methanomicrobiales |
Methanoculleus |
| Methanospirillum |
メタン生成反応はメタン生成古細菌単独で進行するばかりでなく,様々な嫌気微生物との共生系によっても進行する。その中で,最も有名な例はモMethanobacillus omelianskiiモ 6)で,メタン生成古細菌と水素生成真正細菌との水素を介した種間水素転移と呼ばれる栄養共生関係に基づくメタン生成共生系である40)。水田土壌においても,プロピオン酸などからこのような共生系を介して生成される水素が,メタン生成反応に重要な役割を果たしていると考えられている9)。最近,水田土壌でプロピオン酸を分解する共生系の微生物群集を安定同位体プロービング(SIP)法により解析した研究28)が報告されたが,関与する微生物の分離培養が困難なこともあり,群集を構成する微生物についての知見は未だ十分ではない。森田ら(未発表)はプロピオン酸を基質とした培地に水田土壌を接種し,継代培養を行って得られた集積培養液について,DGGE法により培養液中に存在する真正細菌群集を解析した。SyntrophomonasやSyntrophobacterなど,これまでに酪酸やプロピオン酸などの低級脂肪酸を分解するメタン生成共生系で知られている水素生成真正細菌群40)に近縁な配列は得られなかったものの,Bacteroidetesに属するProteiniphilum acetatigenesに近縁な配列が得られた。P. acetatigenesは,プロピオン酸を分解する, Syntrophobacter sulfatireduenceとMethanobacterium formicicumからなるメタン生成共生系から得られた菌株で,自身はプロピオン酸の分解もメタンの生成も行わないが,この共生系に共存することにより,プロピオン酸分解とメタン生成を促進することが明らかにされている7)。DNA解析のみからの推定であるため,さらに研究が必要であるが,水田土壌のプロピオン酸分解共生系においても同様の共生的ともいえる相互作用が働いている可能性を示唆する結果である。これまで,共生では二者生物間の関係が主に研究されてきたが,このような三者の微生物からなる共生的な相互作用が,メタン生成共生系のみならず,水田土壌の様々な代謝に関与しているのかもしれない。
窒素代謝とメタン生成という,水田土壌では古くから多くの研究が行われてきた代謝過程を中心に述べたが,鍵となる微生物群や反応に関わる相互作用について,亜硝酸酸化・脱窒やメタン生成共生系の例のように,不明な点もまだ多く残されている。加えて,嫌気的アンモニア酸化46)や硫酸イオン52)及び硝酸イオン37)の還元と共役した嫌気的メタン酸化など新たな反応経路や共生系が近年になって他の嫌気生態系で見つかっており,水田土壌でもこれらの微生物反応が行われているか,また,どのような微生物群が関わっているのか興味深い。今後の研究の進展に期待したい。分子生態学的手法の発達により微生物群集構造の解析はかつてより容易に行えるようになった。一方で,土壌中における機能や微生物間の相互作用を明らかにするためには,上記のプロテアーゼ生産菌やRice
cluster Iに属するメタン生成古細菌,あるいは畑下層土における低栄養性脱窒菌などのように,菌の分離・培養を併せて行っていく必要があろう。
文 献
1)浅川晋(1998)メタン生成古細菌の特徴と水田土壌における生態,土と微生物,52,33〜39
2) 浅川晋(2005)水田における物質循環,土壌サイエンス入門,三枝正彦・木村眞人編,p.74〜84,文永堂,東京
3) 浅沼修一・齋藤雅典(2003)窒素はどこからきたのか?-窒素固定研究の歩み-,新・土の微生物(10)研究の歩みと展望,日本土壌微生物学会編,p.171〜191,博友社,東京
4) 浅利直子・石原利恵・中嶋泰則・木村眞人・浅川晋(2005)水田土壌表層部の稲わらに生息するシアノバクテリアの群集構造の解析,第21回日本微生物生態学会講演要旨集,p.143
5) Baker G and Tiedje JM (2003) Nitric oxide reductase (norB) genes
from pure cultures and environmental samples. Appl. Environ. Microbiol.,
69, 3476-3483
6) Bryant MP, Wolin EA, Wolin MJ, Wolfe RS (1967) Methanobacillus omelianskii,
a symbiotic association of two species of bacteria. Arch. Mikrobiol., 59,
20-31
7) Chen S, Dong X (2005) Proteiniphilum acetatigenes gen. nov.,
sp. nov., from a UASB reactor treating brewery wastewater. Int. J. Syst.
Evol. Microbiol., 55, 2257-2261
8) Chèneby D, Hallet S, Mondon M, Martin-Laurent F, Germon JC, Philippot
L (2003) Genetic characterization of the nitrate reducing community based
on narG nucleotide sequence analysis. Microb. Ecol., 46, 113-121
9) Conrad R, Babbel M (1989) Effect of dilution on methanogenesis, hydrogen
turnover and interspecies hydrogen transfer in anoxic paddy soil. FEMS Microbiol.
Ecol., 62, 21-28
10) Dianou D, Miyaki T, Asakawa S, Morii H, Nagaoka K, Oyaizu H, Matsumoto
S (2001) Methanoculleus chikugoensis sp. nov., a novel methanogenic
archaeon isolated from paddy field soil in Japan, and DNA-DNA hybridization
among Methanoculleus species. Int. J. Syst. Evol. Microbiol., 51,
1663-1669
11) Großkopf R, Stubner S, Liesack W (1998) Novel euryarchaeotal lineage
detected on rice roots and in the anoxic bulk soil of flooded rice microcosms.
Appl. Envirion. Microbiol., 64, 4983-4989
12) Hashimoto T, Whang K-S, Nagaoka K (2006) A qualitative evaluation and
phylogenetic characterization of oligotrophic denitrifying bacteria harbored
in subsurface upland soil using improved culturability. Biol. Fertil. Soils,
42, 179-185
13) Hayano K (1993) Protease activities in paddy field soil: Origin and
some properties. Soil Sci. Plant Nutr., 39, 539-546
14) 早野恒一(1995)耕地土壌の窒素地力とプロテアーゼ,化学と生物,33,173〜180
15) 早野恒一(1997)植物の窒素,リン栄養と微生物,新・土の微生物(2)植物の生育と微生物,土壌微生物研究会編,p.133〜165,博友社,東京
16) Hayano K and Watanabe K (1990) Characterization of extracellular protease
in paddy field soils. Proceeding of 14th International Congress of Soil
Science, Vol. III. p.270-271
17) 早津雅仁(2002)土壌微生物と物質循環2.硝酸性窒素の動態・処理と土壌微生物,土肥誌,73,323〜329
18) Ikenaga M, Asakawa S, Muraoka Y and Kimura M (2003) Phylogenetic study
on CTO primer-amplyfied ammonia-oxidizing bacteria and b-Proteobacteria
associated with rice roots grown in a flooded paddy soil. Soil Sci. Plant
Nutr., 49, 719-727
19) 井町寛之・酒井早苗(2006)難培養性メタン生成古細菌Rice Cluster Iの分離と特性,土と微生物,60,85〜89
20) Jia Z, Sugano A, Asari N, Tun CC, Ishihara R, Asakawa S and Kimura
M (2005) Molecular characterization of methane oxidizing bacteria inhabiting
rice straw incorporated into paddy field. 土肥要旨集,51,40
21) Jia Z, Kikuchi H, Watanabe T, Asakawa S and Kimura M (submitted for
publication) Molecular identification of methane-oxidizing bacteria in rice
field soil. Biol. Fertil. Soils
22) 菊地弘泰・渡邉健史・Jia Z・木村眞人・浅川晋(2005)水田作土中の真正細菌群集構造のPCR-DGGE法による解析,土肥要旨集,51,41
23) 金澤晋二郎(1982)水田土壌の生物,水田土壌学,山根一郎編,p.233〜279,農文協,東京
24) Kimura M (2005) Populations, community composition and biomass of aquatic
organisms in the floodwater of rice fields and effects of field management.
Soil Sci. Plant Nutr., 51, 159-181
25) Kimura M and Asakawa S (2006) Comparison of community structures of
microbiota at main habitats in rice field ecosystems based on phospholipid
fatty acid analysis. Biol. Fertil. Soils, 43, 20-29
26) Kowalchuk GA and Stephen JR (2001) Ammonia-oxidizing bacteria: a model
for molecular microbial ecology. Annu. Rev. Microbiol., 55, 485-529
27) 小山雄生(1975)15N利用による水田土壌窒素肥沃度測定の実際と生産力,土肥誌,46,260〜269
28) Lueders T, Pommerenke B, Friedrich MW (2004) Stable-isotopic probing
of microorganisms thriving at thermodynamic limits: syntrophic propionate
oxidation in flooded soil. Appl. Environ. Microbiol., 70, 5778-5786
29) Matsuyama T, Nakajima Y, Matsuya K, Ikenaga M, Asakawa S, Kimura M
(2007) Bacterial community in plant residues in a Japanese paddy field estimated
by RFLP and DGGE analyses. Soil Biol. Biochem., 39, 463-472
30) 陽捷行(1998)農林生態系の環境保全機能と環境影響,環境保全と農林業,陽捷行編著,p.1〜17,朝倉書店,東京
31) 水上里美・武田潔・赤田辰治・藤田隆(2001)水田土壌の糸状性酢酸利用性メタン生成菌の分布と特徴,土肥誌,72,633〜641
32) Mizukami S, Takeda K, Akada S, Fujita T (2006) Isolation and characteristics
of Methanosaeta in paddy field soils. Biosci. Biotechnol. Biochem.,
70, 828-835
33) Murase J, Itoh K, Kano M and Kimura M (2003) Molecular analysis of
b-proteobacterial ammonia oxidizer populations in surface layers of a submerged
paddy soil microcosm. Soil Sci. Plant Nutr., 49, 909-913
34) 日本土壌肥料学会編(1990)水田土壌の窒素無機化と施肥,博友社,181p,東京
35) Niswati A, Murase J, Asakawa S and Kimura M (2004) Analysis of communities
of ammonia oxidizers, methanotrophs, and methanogens associated with microcrustaceans
in the floodwater of a rice field microcosm. Soil Sci. Plant Nutr., 50,
447-455
36) 小野信一・古賀汎(1984)水田土壌表層における窒素の自然集積とラン藻による窒素固定,土肥誌,55,465〜470
37) Rafhoebarsing AA, Pol A, van de Pas-Schoonen KT, Smolders AJP, Ettwig
KF, Rijpstra WIC, Schouten S, Damst・JSS, Op den Camp HJM, Jetten MSM, Strous
M (2006) A microbial consortium couples anaerobic methane oxidation to denitrification.
Nature, 440, 918-921
38) Roger PA, Ladha JK (1992) Biological N2 fixation in wetland
rice fields: estimation and contribution to nitrogen balance. Plant Soil,
141, 41-55
39) 芝垣武俊・中嶋泰則・松家一夫・浅川晋・村瀬潤・木村眞人(2004)田面水中のアンモニア酸化細菌に関する研究:分子生物学的手法による田面水からのアンモニア酸化菌の検出の試み,土肥要旨集,50,293
40) Schink B (1997) Energetics of syntrophic cooperation in methanogenic
degradation. Microbiol. Mol. Biol. Rev., 61, 262-280
41) Schramm A (2003) In situ analysis of structure and activity of the
nitrifying community in biofilms, aggregates and sediments. Geomicrobiol.
J., 20, 313-333
42) 塩入松三郎・青峰重範(1937)水田状態の土壌中におけるアンモニア態窒素の消長について,土肥誌,11,389〜392
43) 塩入松三郎・青峰重範(1938)湛水状態の土壌中における窒素の形態変化,土肥誌,12,406〜410
44) 庄子貞雄・前忠彦(1984)無機養分と水の動態,作物の生態生理,佐藤庚・山崎耕宇・庄子貞雄・前忠彦・秋田重誠・中條博良・玖村敦彦・渡辺和之著,p.97〜171,文永堂,東京
45) Song T, M罫tensson L, Eriksso T, Zheng W, Rasmussen U (2005) Biodiversity
and seasonal variation of the cyanobacterial assemblage in a rice paddy
field in Fujian, China. FEMS Microbiol. Ecol., 54, 131-140
46) Stous M, Fuerst JA, Kramer EHM, Logemann S, Muyzer G, van de Pas-Schoonen
KT, Webb R, Kuenen JG, Jetten MSM (1999) Missing lithotroph identified as
new planctomycete. Nature, 400, 446-449
47) Teske A, Elm E, Regan JM, Toze S, Rittmann BE and Stahl DA (1994) Evolutionary
relationships among ammonia- and nitrite-oxidizing bacteria. J.Bacteriol.,
176, 6623-6630
48) Throbäck IN, Enwall K, Jarvis Å, Hallin S (2004) Reasseing PCR primers
targeting nirS, nirK and nosZ genes for community surveys
of denitrifying bacteria with DGGE. FEMS Microbiol. Ecol., 49, 401-417
49) Tonouchi A (2002) Isolation and characterization of a mobile hydrogenotrophic
methanogen from rice paddy field soil in Japan. FEMS Microbiol. Lett., 208,
239-243
50) 鳥山和伸・関矢信一郎(2006)水稲の施肥,肥料の事典,尾和尚人・木村眞人・越野正義・三枝正彦・但野利秋・長谷川功・吉羽雅昭編,p.222〜230,朝倉書店,東京
51) 辻村茂男(2000)土壌藻類の働きと利用,新・土の微生物(7)生態的に見た土の原生動物・藻類,日本土壌微生物学会編,p.127〜158,博友社,東京
52) Valentine DL, Reeburgh WS (2000) New perspective on anaerobic methane
oxidation. Environ. Microbiol., 2, 477-484
53) Watanabe A (1959) Distribution of nitrogen-fixing blue-green algae
in various areas of south and east asia. J. Gen. Appl. Microbiol., 5,
21-29
54) Watanabe A, Yamamoto Y (1971) Algal nitrogen fixation in the tropics.
Plant Soil, Special Volume, 403-423
55) 渡辺巌・Roger PA (1983)水田土壌の窒素固定微生物の動態,微生物の生態11変動と制御をめぐって,微生物生態研究会編,p.31〜53,学会出版センター,東京
56) Watanabe K and Hayano K (1993a) Distribution and identification of
proteolytic Bacillus spp. in paddy field soil under rice cultivation.
Can. J. Microbiol., 39, 674-680
57) Watanabe K and Hayano K (1993b) Source of soil protease in paddy fields.
Can. J. Microbiol., 39, 1035-1040
58) Watanabe K and Hayano K (1994) Source of soil protease based on the
splitting sites of a polypeptide. Soil Sci. Plant Nutr., 40,697-701
59) Watanabe K, Asakawa S and Hayano K (1994) Evaluation of extracellular
protease activities of soil bacteria. Soil Biol. Biochem., 26, 479-482
60) Watanabe K and Hayano K (1995) Seasonal variation of soil protease
activities and their relation to proteolytic bacteria and Bacillus
spp. in paddy field soil. Soil Biol. Biochem., 27, 197-203
61) Watanabe K and Hayano K (1996) Seasonal variation in extracted proteases
and relationship to overall soil protease and exchangeable ammonia in paddy
soils. Biol. Fertil. Soils, 21, 89-94
62) Watanabe K and Hayano K (1996) Mechanisms of production of soil protease
by proteolytic Bacillus subtilis in wetland rice soil. Biol. Fertil.
Soils, 21, 109-113
63) 渡邊克二・早野恒一(1996)土壌中のプロテアーゼ生産微生物,土と微生物,47,9〜22
64) Watanabe T, Kimura M, Asakawa S (2006) Community structure of methanogenic
archaea in paddy field soil under double cropping (rice-wheat). Soil Biol.
Biochem., 38, 1264-1274
65) 山本勇(2000)脱窒を担う細菌と遺伝子群,化学と生物,38,301〜308
66) Zumft WG (1997) Cell biology and molecular basis of denitrification.
Microbiol. Mol. Biol. Rev., 61, 533-616
土壌生物化学のページへ